Citation: Hossein Mozdarani, Sohail Mozdarani. De novo cytogenetic alterations in spermatozoa of subfertile males might be due to genome instability associated with idiopathic male infertility: Experimental evidences and Review of the literature[J]. AIMS Genetics, 2016, 3(4): 219-238. doi: 10.3934/genet.2016.4.219

| [1] |
Sharpe RM (2012) Sperm counts and fertility in men: a rocky road ahead. EMBO Rep 13: 398-403. doi: 10.1038/embor.2012.50

|
| [2] | Rolland M, Le Moal J, Wagner V, et al. (2013) Decline in semen concentration and morphology in a sample of 26 609 men close to general population between 1989 and 2005 in France. Hum Reprod 28: 462-470. |
| [3] | McLachlan RI, de Kretser DM (2001) Male infertility: the case for continued research. Med J 174: 116-117. |
| [4] | Swan SH, Elkin EP, Fenster L (1997) Have sperm densities declined? A reanalysis of global trend data. Environ Health Perspect 11: 1228-1232. |
| [5] | Gaspari L, Paris F, Jandel C, et al. (2011) Prenatal environmental risk factors for genital malformations in a population of 1442 French male newborns: a nested case-control study. Hum Reprod 11: 3155-3162. |
| [6] |
Moller P, Wallin H, Knudsen L (1996) Oxidative stress associated with exercise, psychological stress and lifestyle factors. Chem-Biol Interac 102: 17-36. doi: 10.1016/0009-2797(96)03729-5
|
| [7] |
Fraga C, Motchnik P, Shigenaga M, et al. (1991) Ascorbic acid protects against endogenous oxidative DNA damage in human sperm. Proc Natl Acad Sci USA 88: 11003-11006. doi: 10.1073/pnas.88.24.11003
|
| [8] | Shen HM, Chia SE, Ni ZY, et al. (1997) Detection of oxidative DNA damage in human sperm and the association with cigarette smoking. Reprod Technol 11: 675-680. |
| [9] |
Nili HA, Mozdarani H, Aleyasin A (2009) Correlation of sperm DNA damage with protamine deficiency in Iranian subfertile men. Reprod BioMed Online 18: 479-485. doi: 10.1016/S1472-6483(10)60123-X
|
| [10] | Nili HA, Mozdarani H, Pellestor F (2011) Impact of DNA damage on the frequency of sperm chromosomal aneuploidy in normal and subfertile men. Iranian Biomedical J 15: 122-129. |
| [11] | Lewis SE, Aitken JR, Conner SJ, et al. (2013) The impact of sperm DNA damage in assisted conception and beyond: recent advances in diagnosis and treatment. Reprod Biomed Online 27: 325-337. |
| [12] |
Wright C, Milne S, Leeson H (2014) Sperm DNA damage caused by oxidative stress: modifiable clinical, lifestyle and nutritional factors in male infertility. Reprod Biomed Online 28: 684-703. doi: 10.1016/j.rbmo.2014.02.004
|
| [13] | Agarwal A, Mulgund A, Alshahrani S, et al. (2014) Reactive oxygen species and sperm DNA damage in infertile men presenting with low level leukocytospermia. Reprod Biol Endocrinol 12: 126. |
| [14] | Hassold TJ (1998) Nondisjunction in the human male. Current Topics in Development Biology 37: 383-406. |
| [15] |
Shi Q and Martin RH (2001) Aneuploidy in human spermatozoa: FISH analysis in men with constitutional chromosomal abnormalities, and in infertile men. Reproduction 121: 655-666. doi: 10.1530/rep.0.1210655
|
| [16] |
Collodel G, Capitani S, Baccetti B, et al. (2007) Sperm aneuploidies and low progressive motility. Hum Reprod 22: 1893-1898. doi: 10.1093/humrep/dem099
|
| [17] |
Kirkpatrick G, Ferguson KA, Gao H, et al. (2008) A comparison of sperm aneuploidy rates between infertile men with normal and abnormal karyotypes. Hum Reprod 23: 1679-1683. doi: 10.1093/humrep/den126
|
| [18] | Warburton D, Kline J, Stein Z, et al. (1980) Monosomy X: a chromosomal anomaly associated with young maternal age. Lancet 1: 167-169. |
| [19] | Hook EB (1985) Maternal age, paternal age, and human chromosome abnormality: nature, magnitude, etiology, and mechanisms of effects. Basic Life Science 36: 117-132. |
| [20] | Mozdarani H, Aghdaei F (2001) Cytogenetic analysis of failed fertilized oocytes from Iranian infertile women after in vitro fertilization (IVF) and intracytoplasmic sperm injection (ICSI) procedures. Middle East Fertility Society J 6: 216-225. |
| [21] | Mozdarani H, Sammak L (2001) Chromosomal abnormalities of human preimplantation embryos generated from in vitro fertilization and intracytoplasmic sperm injection. Middle East Fertility Society J 6: 163-168. |
| [22] |
Vegetti W, Van Assche E, Frias A, et al. (2000) Correlation between semen parameters and sperm aneuploidy rates investigated by fluorescence in situ hybridization in infertile men. Hum Reprod 15: 351-365. doi: 10.1093/humrep/15.2.351
|
| [23] | Hristova R, Ko E, Greene C, et al. (2002) Chromosome abnormalities in sperm from infertile men with asthenoteratozoospermia. Biol Reprod 66: 1781-1783. |
| [24] |
Storeng RT, Plachot M, Theophile D, et al. (1998) Incidence of sex chromosome abnormalities in spermatozoa from patients entering an IVF or ICSI protocol. Acta Obstetricia et Gynecologica Scandinavica 77: 191-197. doi: 10.1080/j.1600-0412.1998.770212.x
|
| [25] |
Blanco J, Gabau E, Gomez D, et al. (1998) Chromosome 21 disomy in spermatozoa of the fathers of children with trisomy 21, in a population with a high prevalence of Down syndrome: increased incidence in cases of paternal origin. Am J Hum Genet 63: 1067-1072. doi: 10.1086/302058
|
| [26] |
Martinez-Pasarell O, Marquez C, Coll MD, et al. (1997) Analysis of human sperm-derived pronuclei by three-colour fluorescent in-situ hybridization. Hum Reprod 12: 641-645. doi: 10.1093/humrep/12.4.641
|
| [27] |
Moosani N, Chernos J, Lowry RB, et al. (1999) A 47,XXY fetus resulting from ICSI in a man with an elevated frequency of 24,XY sperm. Hum Reprod 14: 1137-1138. doi: 10.1093/humrep/14.4.1137
|
| [28] |
Bonduelle M, Van Assche E, Jordis H, et al. (2002) Prenatal testing in ICSI pregnancies: incidence of chromosomal abnormalities in 1586 karyotypes and relation to sperm parameters. Hum Reprod 17: 2600-2614. doi: 10.1093/humrep/17.10.2600
|
| [29] |
Tang SS, Gao H, Robinson WP, et al. (2004) An association between sex chromosomal aneuploidy in sperm in an abortus with 45, X of paternal origin: possible transmission of chromosomal abnormalities through ICSI. Hum Reprod 19: 147-151. doi: 10.1093/humrep/deh007
|
| [30] |
Tiepolo L and Zuffardi O (1976) Localization of factors controlling spermatogenesis in the nonfluorescent portion of the human Y chromosome long arm. Hum Genet 34: 119-124. doi: 10.1007/BF00278879
|
| [31] |
Vog P, Edelmann A, Kirsch S, et al. (1996) Human Y chromosome azoospermia factors (AZF) mapped to different subregions in Yq11. Hum Mol Gen 5: 933-943. doi: 10.1093/hmg/5.7.933
|
| [32] |
Kuroda-Kawaguchi T, Skaletsky H, Brown LG, et al. (2001) The AZFc region of the Y chromosome features massive palindromes and uniform recurrent deletions in infertile men. Nat Genet 29: 279-286. doi: 10.1038/ng757
|
| [33] |
Skaletsky H, Kuroda-Kawaguchi T, Minx PJ, et al. (2003) The male-specific region of the human Y chromosome is a mosaic of discrete sequence classes. Nature 423: 825-837. doi: 10.1038/nature01722
|
| [34] | McElreavey K, Krausz C, Bishop CE. (2000) The human Y chromosome and male infertility. Results Probl Cell Differ 28: 2211-2232. |
| [35] |
Van der Ven K, Montag M, Peschka B, Leygraaf J, et al. (1997) Combined cytogenetic and Y chromosome microdeletion screening in males undergoing intracytoplasmic sperm injection. Mol Hum Reprod 3: 699-704. doi: 10.1093/molehr/3.8.699
|
| [36] |
Foresta C, Ferlin A, Garolla A, et al. (1998) High frequency of well-defined Y-chromosome deletions in idiopathic Sertoli cell-only syndrome. Hum Reprod 13: 302-307. doi: 10.1093/humrep/13.2.302
|
| [37] |
Katagiri Y, Neri Q V, Takeuchi T, et al. (2004) Y chromosome assessment and its implications for the development of ICSI children. Reprod Biomed Online 8: 307-318. doi: 10.1016/S1472-6483(10)60911-X
|
| [38] | Foresta C, Bettella A, Moro E, et al. (2001) Sertoli Cell Function in Infertile Patients with and without Microdeletions of the Azoospermia Factors on the Y Chromosome Long Arm. J Clin Endocrinol Metab 86: 2414-2419. |
| [39] | Yu XW, Wei ZT, Jiang YT, et al. (2015) Y chromosome azoospermia factor region microdeletions and transmission characteristics in azoospermic and severe oligozoospermic patients. Int J Clin Exp Med 8: 14634-14646. |
| [40] |
Krausz C and McElreavey K (1999) Y chromosome and male infertility. Front Biosci 4: E1-E8. doi: 10.2741/A399
|
| [41] |
Maurer B, Simoni M (2000) Y chromosome microdeletion screening in infertile men. J Endocrinol Invest 23: 664-670. doi: 10.1007/BF03343791
|
| [42] |
Simoni M, Bakker E, Krausz C (2004) EAA/EMQN best practice guidelines for molecular diagnosis of y-chromosomal microdeletions. State of the ART. Int J Androl 27: 240-249. doi: 10.1111/j.1365-2605.2004.00495.x
|
| [43] |
Dada R, Kumar R, Shamsi MB, et al. (2007) Higher frequency of Yq microdeletions in sperm DNAas compared to DNA isolated from blood. Asian J Androl 9: 720-722. doi: 10.1111/j.1745-7262.2007.00274.x
|
| [44] |
Mozdarani H, Ghoraeian P (2012) Efficient combined FISH and PRINS technique for detection of DAZ microdeletion in human sperm. J Assist Reprod Genet 29: 979-84. doi: 10.1007/s10815-012-9805-z
|
| [45] |
Lewis SE and Agbaje IM (2008) Using the alkaline comet assay in prognostic tests for male infertility and assisted reproductive technology outcomes. Mutagenesis 23: 163-170. doi: 10.1093/mutage/gem052
|
| [46] |
Speit G, Vasquez M, Hartmann A (2009) The comet assay as an indicator test for germ cell genotoxicity. Mutat Res 681: 3-12. doi: 10.1016/j.mrrev.2008.03.005
|
| [47] |
Shahidi M, Mozdarani H, Bryant PE (2007) Radiation sensitivity of leukocytes from healthy individuals and breast cancer patients as measured by the alkaline and neutral comet assay. Cancer Letters 257: 263-273. doi: 10.1016/j.canlet.2007.08.002
|
| [48] |
Rudak E, Jacobs PA, Yanagimachi R (1978) Direct analysis of the chromosome constitution of human spermatozoa. Nature 274: 911-913. doi: 10.1038/274911a0
|
| [49] | Martin RH, Lin CC, Balkan W, et al. (1982) Direct chromosomal analysis of human spermatozoa: preliminary results from 18 normal men. Am J Human Genet 34: 459-468. |
| [50] |
Martin RH, Rademaker AW, Greene C, et al. (2003) A comparison of the frequency of sperm chromosome abnormalities in men with mild, moderate, and severe oligozoospermia. Biol Reprod 69: 535-539. doi: 10.1095/biolreprod.102.015149
|
| [51] | Sun F, Turek P, Greene C, et al. (2007) Abnormal progression through meiosis in men with nonobstructive azoospermia. Fertil Steril 87: 565-571. |
| [52] |
Pellestor F, Imbert I, Andréo B (2002) Rapid chromosome detection by PRINS in human sperm. Am J Med Genet 107: 109-114. doi: 10.1002/ajmg.10101
|
| [53] | World Health Organization (2010) WHO laboratory manual for the examination and processing of human semen. Fifth edition; WHO Press. |
| [54] |
Aitken RJ and Clarkson JS (1988) Significance of reactive oxygen species and antioxidants in defining the efficacy of sperm preparation techniques. J Androl 9: 367-376. doi: 10.1002/j.1939-4640.1988.tb01067.x
|
| [55] | Singh NP (1996) Microgel electrophoresis of DNA. In: Pfeifer GP, editor. Technologies for Detection of DNA Damage and Mutations. New York: Plenum Press Pp: 3-24. |
| [56] |
Jaloszynski P, Kujawski M, Czub-Swierczek M, et al. (1997) Bleomycin-induced DNA damage and its removal in lymphocytes of breast cancer patients studied by comet assay. Mutat Res 385: 223-233. doi: 10.1016/S0921-8777(97)00046-3
|
| [57] |
Pellestor F, Paulasova P, Andréo B, et al. (2006) Multicolor PRINS and multicolor PNA. Cytogenet Genom Res 114: 263-269. doi: 10.1159/000094211
|
| [58] |
Kadandale JS, Wachtel SS, Tunca Y, et al. (2002) Deletion of RBM and DAZ in azoospermia: evaluation by PRINS. Am J Med Genet 107: 105-108. doi: 10.1002/ajmg.10107
|
| [59] |
Gao JL, Nie Y, Ding XP. (2011) Primed in situ labeling for detecting single-copy genes. Genet Mol Res 10: 1884-1890. doi: 10.4238/vol10-3gmr1391
|
| [60] |
Zini A and Libman J (2006) Sperm DNA damage: clinical significance in the era of assisted reproduction. Can Med Assoc J 175: 495-500. doi: 10.1503/cmaj.060218
|
| [61] | Perry MJ (2008) Effects of environmental and occupational pesticide exposure on human sperm: a systematic review. Hum Reprod Updat 14: 233-242. |
| [62] |
Barratt CL, Aitken RJ, Björndahl L, et al. (2010) Sperm DNA: organization, protection and vulnerability: from basic science to clinical applications--a position report. Hum Reprod 25: 824-838. doi: 10.1093/humrep/dep465
|
| [63] | Kodama H, Yamaguchi R, Fukuda J, et al. (1997) Increased oxidative deoxyribonucleic acid damage in the spermatozoa of infertile male patients. Fertil Steril 68: 519-524. |
| [64] |
Enciso M, Sarasa J, Agarwal A, et al. (2009) A two-tailed Comet assay for assessing DNA damage in spermatozoa. Reprod Biomed Online 18: 609-616. doi: 10.1016/S1472-6483(10)60003-X
|
| [65] |
Kumar D, Salian SR, Kalthur G, et al. (2013) Semen Abnormalities, Sperm DNA Damage and Global Hypermethylation in Health Workers Occupationally Exposed to Ionizing Radiation. PLoS ONE 8: e69927. doi: 10.1371/journal.pone.0069927
|
| [66] |
Santos R, Palos-Ladeiro M, Besnard A, et al. (2013) Relationship between DNA damage in sperm after ex vivo exposure and abnormal embryo development in the progeny of the three-spined stickleback. Reprod Toxicol 36: 6-11. doi: 10.1016/j.reprotox.2012.11.004
|
| [67] | De Iuliis GN, Newey RJ, King BV, et al. (2014) Mobile Phone Radiation Induces Reactive Oxygen Species Production and DNA Damage in Human Spermatozoa In Vitro. PLOS ONE |
| [68] | Aitken RJ, Ryan AL, Curry BJ, et al. (2003) Multiple forms of redox activity in populations of human spermatozoa. Mol Hum Reprod 9: 645-661. |
| [69] |
Pathak D, Premi S, Srivastava J, et al. (2006) Genomic instability of the DYZ1 repeat in patient with Y chromosome anomalies and males exposed to natural background radiation. DNA Research 13: 103-109. doi: 10.1093/dnares/dsl002
|
| [70] |
Premi S, Srivastava J, Chandy SP, et al. (2006) Tandem duplication and copy number polymorphism of the SRY gene in patients with sex chromosome anomalies and males exposed to natural background radiation. Mol Hum Reprod 12: 113-121. doi: 10.1093/molehr/gal012
|
| [71] |
Premi S, Srivastava J, Chandy SP, et al. (2007) AZFc somatic microdeletions and copy number polymorophism of the DAZ genes in human males exposed to natural background radiation. Hum Genet 121: 337-346. doi: 10.1007/s00439-006-0318-7
|
| [72] |
Arruda JT, Silva DM, Silva CC, et al. (2008) Homologous recombination between HERVs causes duplications in the AZFa region of men accidentally exposed to cesium-137 in Goiânia. Genet Mol Res 7: 1063-1069. doi: 10.4238/vol7-4gmr492
|
| [73] |
Arruda JT (2009) Occurrence of mutations in loci linked to Y chromosome in the offspring born to individuals exposed to ionizing radiation. Genet Mol Res 8: 938. doi: 10.4238/vol8-3ta021
|
| [74] | Moghbeli-Nejad S, Mozdarani H, Behmanesh M, et al. (2012) Genome instability in AZFc region on Y chromosomein leukocytes of fertile and infertile individuals following exposure to gamma radiation. J Assist Reprod Genet 29: 53-61. |
| [75] | Gomez E, Irvine DS, Aitken RJ (1998) Evaluation of a spectrophotometric assay for the measurement of malondialdehyde and 4-hydroxyalkenals in human spermatozoa: relationships with semen quality and sperm function. Int J Androl 21: 81-94. |
| [76] | Simoni M, Gromoll J, Dworniczak B, et al. (2010) Screening for deletions of the Y chromosome involving the DAZ (Deleted in AZoospermia) gene in azoospermia and severe oligozoospermia. Fertil Steril 67: 1753-1756. |
| [77] |
Mozdarani H and Salimi M (2006) Numerical chromosome abnormalities in 8-cell embryos generated from gamma-irradiated male mice in the absence and presence of vitamin E. Int J Radiat Biol 82: 817-822. doi: 10.1080/09553000600973343
|
| [78] |
Mozdarani H and Nazari E. (2009) Cytogenetic damage in preimplantation mouse embryos generated after paternal and parental gamma-irradiation and the influence of vitamin C. Reproduction 137: 35-43. doi: 10.1530/REP-08-0073
|
| [79] | Colpi GM, Contalbi GF, Nerva F, et al. (2004) Testicular function following chemo-radiotherapy. Eur J Obstet Gynecol Reprod Biol 113S: S2-S6. |
| [80] |
Perrin A, Caer E, Oliver-Bonet M, et al. (2009) DNA fragmentation and meiotic segregation in sperm of carriers of a chromosomal structural abnormality. Fertil Steril 92: 583-589. doi: 10.1016/j.fertnstert.2008.06.052
|
| [81] |
Aitken RJ, De Iuliis GN (2010) On the possible origins of DNA damage in human spermatozoa. Molecular Hum Reprod 16: 3-13. doi: 10.1093/molehr/gap059
|
| [82] | Mikamo K, Kamiguchi Y, Tateno H (1991) The interspecif in vitro fertilization system to measure human sperm chromosomal damage. Prog Clin Biol Res 372: 531-542. |
| [83] |
Martin RH, Ko E, Rademaker A (1991) Distribution of aneuploidy in human gametes: Comparison between human sperm and oocytes. Am J Med Genet 39: 321-331. doi: 10.1002/ajmg.1320390315
|
| [84] |
Wyrobek AJ, Robbins WA, Mehraien Y, et al. (1994) Detection of sex chromosomal aneuploidies X-X, Y-Y, and X-Y in human sperm using two-chromosome fluorescence in situ hybridization. Am J Med Genet 53: 1-7. doi: 10.1002/ajmg.1320530102
|
| [85] |
Robbins WA, Meistrich ML, Cassel MJ, et al. (1997) Chemotherapy induces transient sex chromosomal and autosomal aneuploidy in human sperm. Nat Genet 16: 74-78. doi: 10.1038/ng0597-74
|
| [86] | Wyrobek AJ, Rubes J, Cassel M, et al. (1995) Smokers produce more aneuploid sperm than non-smokers. Am J Hum Genet 57: A737. |
| [87] |
Ribas G, Frenzilli G, Barale R, et al. (1995) Herbicide-induced DNA damage in human lymphocytes evaluated by the single cell gel electrophoresis (SCGE) assay. Mutat Res 344: 41-54. doi: 10.1016/0165-1218(95)90037-3
|
| [88] | Marchetti F, Lowe X, Moore DH II, et al. (1996) Paternally inheritied chromosomal structure aberrations detected in mouse firstcleaving zygote metaphases by multicolour fluorescence in situ hybridization painting. Chromosome Res 4: 604-613. |
| [89] | Rupa DS, Schuler M, Eastmond DA (1997) Detection of hyperploidy and breakage affecting the 1cen-1q12 region of cultured interphase human lymphocytes treated with various genotoxic agents. Environ Mol Mutagen 29: 161-167. |
| [90] |
Kinakin B, Rademaker A, Martin R (1997) Paternal age effect of YY aneuploidy in human sperm, as assessed by fluorescence in situ hybridization. Cytogenet Cell Genet 78: 116-119. doi: 10.1159/000134641
|
| [91] | Martin RH, Spriggs E, Ko E, Rademaker AW. (1995)The relationship between paternal age, sex ratios, and aneuploidy frequencies in human sperm, as assessed by multicolor FISH. Am J Med Genet 57: 1393-1399. |
| [92] |
Downie SE, Flaherty SP, Swann NJ, et al. (1997) Estimation of aneuploidy for chromosomes 3, 7, 16, X and Y in spermatozoa from 10 normospermic men using fluorescence in-situ hybridization. Mol Hum Reprod 3: 815-819. doi: 10.1093/molehr/3.9.815
|
| [93] | Egozcue J, Blanco J, Vidal F (1997) Chromosome studies in human sperm nuclei using fluorescence in-situ hybridization (FISH). Hum Reprod Update 3: 441-452. |
| [94] |
Vidal F, Blanco J, Egozcue J (2001) Chromosomal abnormalities in sperm. Mol Cell Endocrinol 183: S51-S54. doi: 10.1016/S0303-7207(01)00579-2
|
| [95] |
Martin RH, Spriggs E, Rademaker AW. (1996) Multicolor Fluorescence In Situ Hybridization Analysis of Aneuploidy and Diploidy Frequencies in 225 846 Sperm from 10 Normal Men. Biol Reprod 54: 394-398. doi: 10.1095/biolreprod54.2.394
|
| [96] |
Ohashi Y, Mihari N, Honda H, et al. (2001) High frequency of XY disomy in spermatozoa of severe oligospermic men. Hum Reprod 16: 703-708. doi: 10.1093/humrep/16.4.703
|
| [97] |
Lynn A, Koehler KE, Judis L, et al. (2002) Covariation of synaptonemal complex length and mammalian meiotic exchange rates. Science 296: 2222-2225. doi: 10.1126/science.1071220
|
| [98] |
Sun F, Trpkov K, Rademaker A, et al. (2005) Variation in meiotic recombination frequencies among human males. Hum Genet 116: 172-178. doi: 10.1007/s00439-004-1215-6
|
| [99] |
Ferguson KA, Chan Wong EA, Chow V, et al. (2007) Abnormal meiotic recombination in infertile men and its association with sperm aneuploidy. Hum Mol Genet 16: 2870-2879. doi: 10.1093/hmg/ddm246
|
| [100] | Martin RH and Rademaker AW (1999) Nondisjunction in human sperm: Comparison of frequencies in acrocentric chromosomes. Cytogenet Cell Genet 86: 43-45. |
| [101] |
Gonsalves J, Sun F, Schlegel PN, et al. (2004) Defective recombination in infertile men. Hum Mol Genet 13: 2875-2883. doi: 10.1093/hmg/ddh302
|
| [102] |
Gianaroli L, Magli MC, Cavallini G, et al. (2005) Frequency of aneuploidy in sperm from patients with extremely severe male factor infertility. Hum Reprod 20: 2140-2152. doi: 10.1093/humrep/dei033
|
| [103] | Baccetti B, Bruni E, Capitani S, et al. (2006) Studies on varicocele III. Ultrastructural sperm evaluation and 18, X and Y aneuploidies. J Androl 27: 94-101. |
| [104] | Templado C, Hoang T, Greene C, et al. (2006) Aneuploid spermatozoa in infertile men: teratozoospermia. Mol Reprod Dev 61: 200-204. |
| [105] |
Machev N, Gosset P, Viville S (2005) Chromosome abnormalities in sperm from infertile men with normal somatic karyotypes: teratozoospermia. Cytogenet Genome Res 111: 352-357. doi: 10.1159/000086910
|
| [106] |
Miharu N (2005) Chromosome abnormalities in sperm from infertile men with normal somatic karyotypes: oligozoospermia. Cytogenet Genome Res 111: 347-351. doi: 10.1159/000086909
|
| [107] |
Ghoraeian P, Mozdarani H, Aleyasin A, et al. (2013) Frequency of sex chromosomal disomy in spermatozoa of normal and oligozoospermic Iranian patients and its effects on fertilization and implantation rates after ICSI. Andrologia 45: 46-55. doi: 10.1111/j.1439-0272.2012.01309.x
|
| [108] |
Reijo R, Lee TY, Salo P, et al. (1995) Diverse spermatogenic defects in humans caused by Y chromosome deletions encompassing a novel RNA-binding protein gene. Nat Genet 10: 383-393. doi: 10.1038/ng0895-383
|
| [109] |
Habermann B, Mi HF, Edelmann A, et al. (1998) DAZ (Deleted in AZospermia) genes encode proteins located in human late spermatids and in sperm tails. Hum Reproduction 13: 363-369. doi: 10.1093/humrep/13.2.363
|
| [110] |
Kent-First MG, Kol S, Muallem A, et al. (1996) The incidence and possible relevance of Y-linked microdeletions in babies born after intracytoplasmic sperm injection and their infertile fathers. Mol Hum Reprod 2: 943-950. doi: 10.1093/molehr/2.12.943
|
| [111] | Kent-First M, Muallem A, Shultz J, et al. (1999) Defining regions of the Y-chromosome responsible for male infertility and identification of a fourth AZF region (AZFd) by Y-chromosome microdeletion detection. Mol Reprod Dev 53: 27-41. |
| [112] |
Samli H, Murat Samli M, Solak M. (2006) Natural transmission of AZFb Y-chromosomal microdeletion from father to his three sons. Arch Androl 52: 423-426. doi: 10.1080/01485010600822655
|
| [113] | Sharlip ID, Jarow JP, Belker AM, et al. (2002) Best practice policies for male infertility. Fertil Steril 5: 873-882. |
| [114] |
Simoni S, Kamischke A, Nieschlag E (1998) Significance of the molecular diagnosis of Y chromosomal microdeletions in the diagnostic workup of male infertility. Hum Reprod 13: 1764-1768. doi: 10.1093/humrep/13.7.1764
|
| [115] |
Koch JE, Kolvraa S, Petersen KB, et al. (1989) Oligonucleotide-priming methods for the chromosome-specific labeling of alpha satellite DNA in situ. Chromosoma 98: 259-65. doi: 10.1007/BF00327311
|
| [116] |
Dai RL, Sun LK, Yang X, et al. (2012) Expansion and de novo occurrence of Y chromosome microdeletions occurring via natural vertical transmission in northeastern China. J Int Med Res 40: 1182-1191. doi: 10.1177/147323001204000339
|
| [117] |
Poongothai JS and Manonayaki S (2008) Y chromosome microdeletions in sperm DNA of infertile patients from Tamil Nadu, south India. Indian Journal of Urology 24: 480-485. doi: 10.4103/0970-1591.44252
|
| [118] |
Writzl K, Zorn B, Peterlin B (2005) Copy number of DAZ genes in infertile men. Fertil Steril 84: 1522-1525. doi: 10.1016/j.fertnstert.2005.06.021
|
| [119] |
Premi S, Srivastava J, Chandy SP, et al. (2007) AZFc somatic microdeletions and copy number polymorphism of the DAZ genes in human males exposed to natural background radiation. Hum Genet 121: 337-346. doi: 10.1007/s00439-006-0318-7
|
| [120] | Zhou DD, Hao JL, Guo KM, et al. (2016) Sperm quality and DNA damage in men from Jilin Province, China, who are occupationally exposed to ionizing radiation. Genet Mol Res 15: gmr.15018078. |
| [121] | Zhang YS, Dai RL, Wang RX, et al. (2014) Azoospermia factor microdeletions: occurrence in infertile men with azoospermia and severe oligozoospermia from China. Andrologia 46: 535-540. |
| [122] |
Le Bourhis C, Siffroi JP, McElreavey K, et al. (2000) Y chromosome microdeletions and germinal mosaicism in infertile males. Mol Hum Reprod 6: 688-693. doi: 10.1093/molehr/6.8.688
|
| [123] |
De Vries JW, Repping S, Oates R, et al. (2001) Absence of deleted in azoospermia (DAZ) genes in spermatozoa of infertile men with somatic DAZ deletions. Fertil Steril 75: 476-479. doi: 10.1016/S0015-0282(00)01758-1
|
| [124] | Tharapel AT, Wachtel SS (2006) PRINS for mapping single-copy genes. Methods Mol Biol 338: 59-67. |
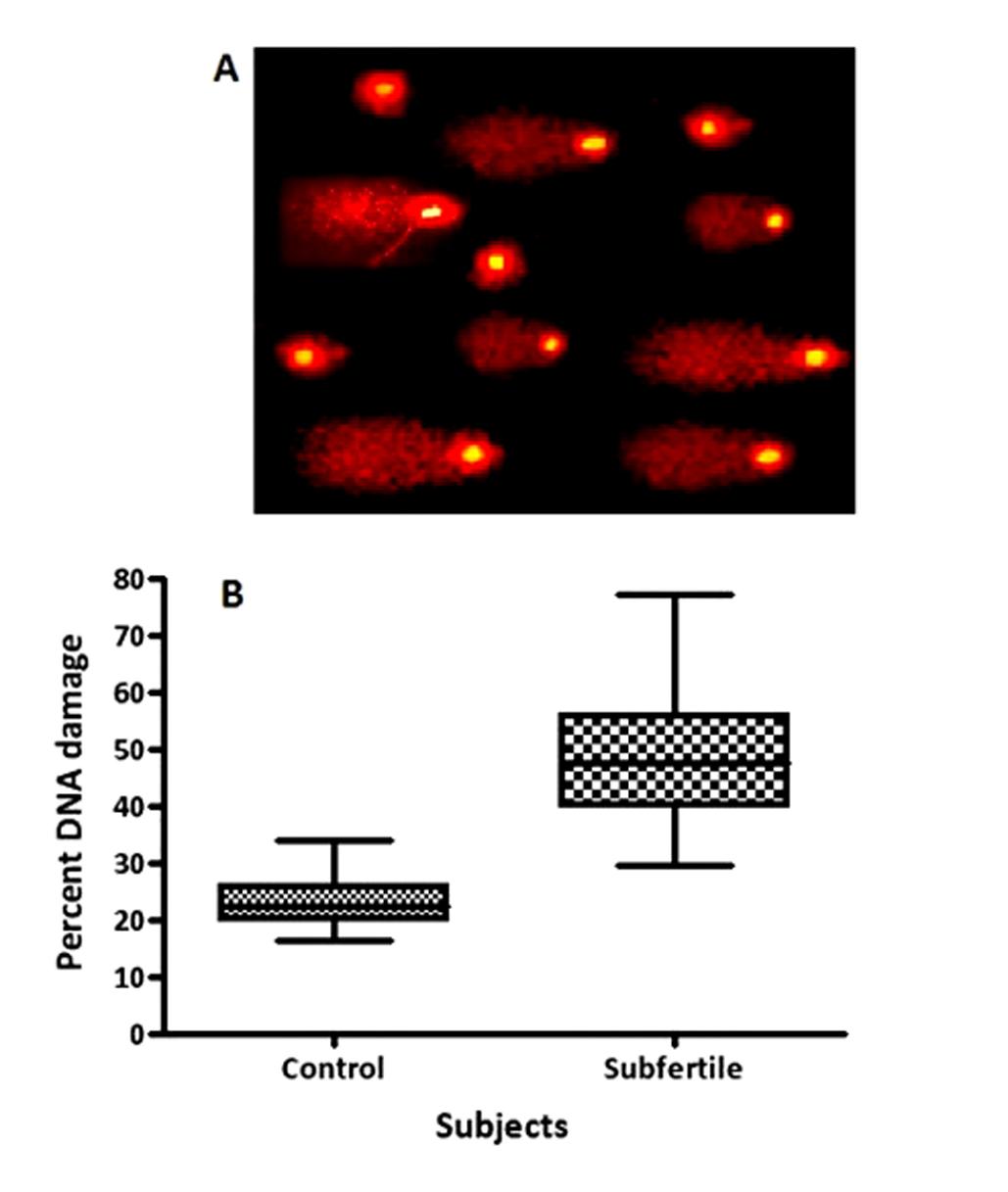
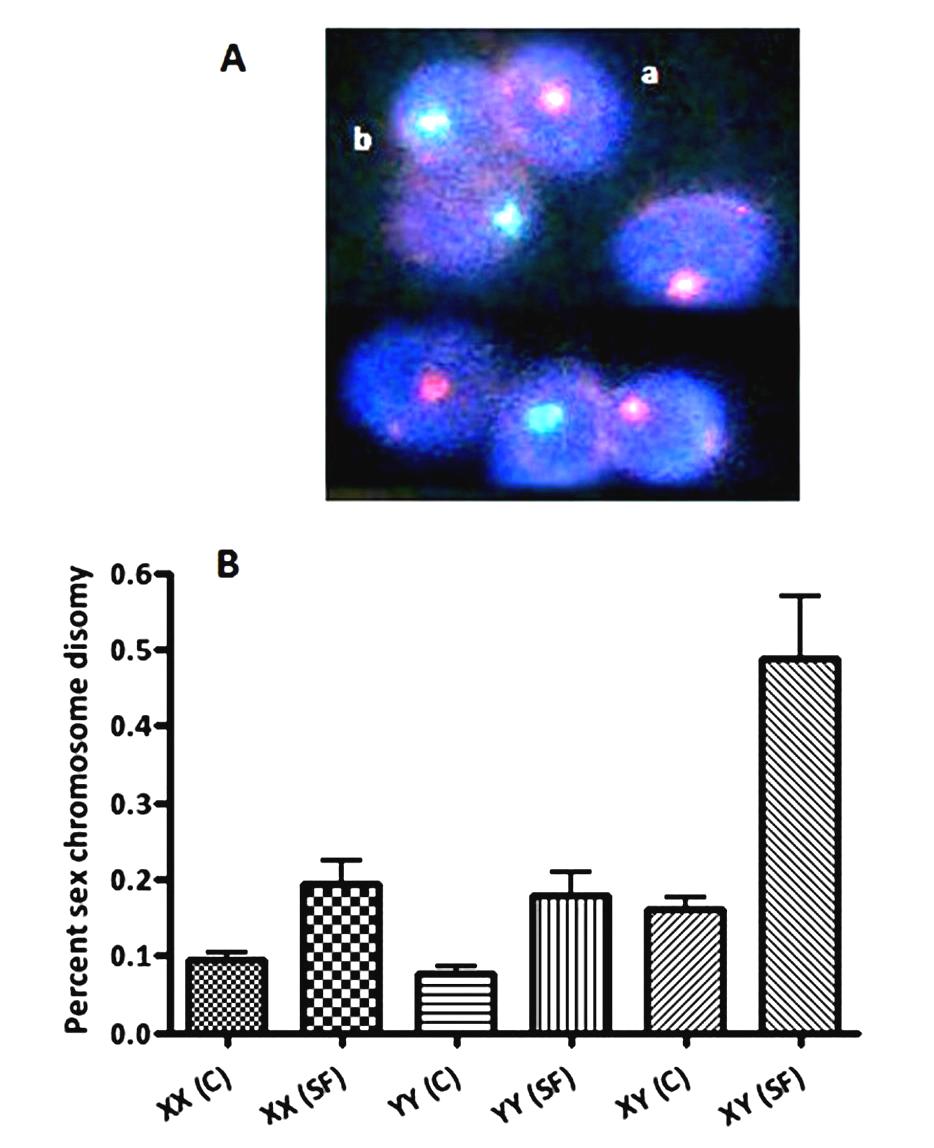
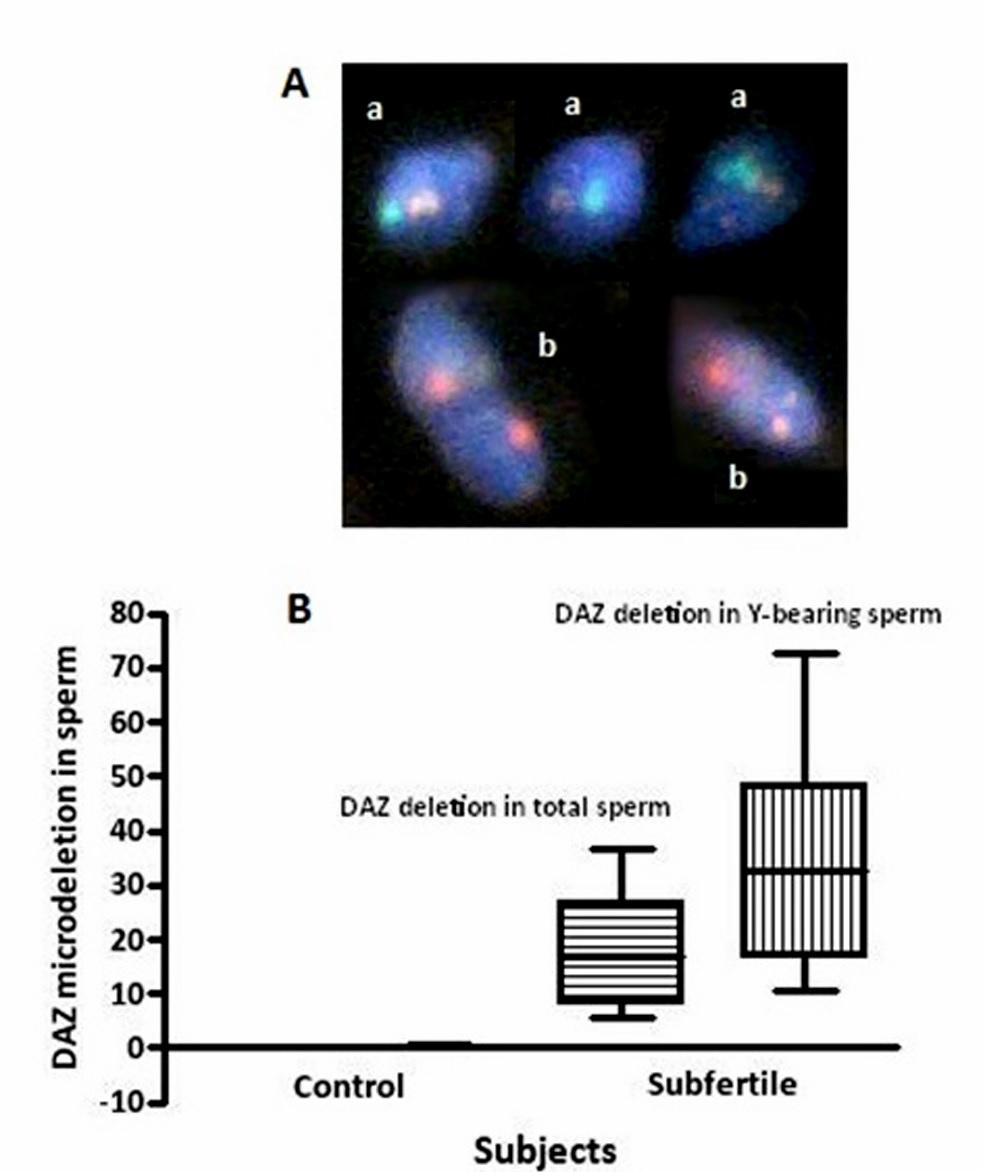
Figures(3) / Tables(1)

Hossein Mozdarani, Sohail Mozdarani. De novo cytogenetic alterations in spermatozoa of subfertile males might be due to genome instability associated with idiopathic male infertility: Experimental evidences and Review of the literature







 DownLoad:
DownLoad: