Citation: Pekka Peura, Patrik Sjöholm. Sustainable or Distributed Energy—or both? Clarifying the Basic Concepts of Reforming the Energy Sector[J]. AIMS Energy, 2015, 3(2): 241-254. doi: 10.3934/energy.2015.2.241

| [1] |
Peura P, Hyttinen T (2011) The potential and economics of bioenergy in Finland. J Cleaner Production 19: 927-945. doi: 10.1016/j.jclepro.2011.02.009

|
| [2] | Peura P (2013a) From Malthus to Sustainable Energy—Theoretical orientations to reforming the energy sector. Renew Sust Energ Rev 19(C): 309-327. |
| [3] | Peura P (2013b) From unlimited growth to sustainable energy. The origins of operational patterns by means of societal selection. Acta Wasaensia 279. 204 p. |
| [4] |
Evrendilek F, Ertekin C (2003) Assessing the potential of renewable energy sources in Turkey. Renew Energ 28: 2303-2315. doi: 10.1016/S0960-1481(03)00138-1
|
| [5] | Ragwitz M, Schleich J, Huber C, et al. (2005) Analyses of the EU renewable energy sources' evolution up to 2020 (FORRES 2020). Stuttgart: Fraunhofer IRB Verlag. |
| [6] |
Lewandowski I, Weger J, van Hooijdonk A, et al. (2006) The potential biomass for energy production in the Czech Republic. Biomass Bioenergy 30: 405-421. doi: 10.1016/j.biombioe.2005.11.020
|
| [7] |
Božičević Vrhovčak M, Tomsic Z, Debrecin N (2006) Potential and use of renewable energy sources in Croatia. Renew Energ 31:1867-1872. doi: 10.1016/j.renene.2005.09.021
|
| [8] |
Schneider DR, Duic N, Bogdan Z (2007) Mapping the potential for decentralized energy generation based on renewable energy sources in the Republic of Croatia. Energy 32: 1731-1744. doi: 10.1016/j.energy.2006.12.003
|
| [9] |
van Dam J, Faaij APC, Lewandowski I, Fischer G (2007) Biomass production in Central and Eastern Europe under different scenarios. Biomass Bioenergy 31: 345-366. doi: 10.1016/j.biombioe.2006.10.001
|
| [10] |
Verbruggen A, Fischedick M, Moomaw W, et al. (2010) Renewable energy costs, potentials, barriers: Conceptual issues. Energy Policy 38: 850-861. doi: 10.1016/j.enpol.2009.10.036
|
| [11] |
Smeets EMW, Faaij AP (2007) Bioenergy potentials from forestry in 2050. Climatic Change 81: 353-390. doi: 10.1007/s10584-006-9163-x
|
| [12] |
Resch G, Held A, Faber T, et al. (2008) Potentials and prospects for renewable energies at global scale. Energy Policy 36: 4048-4056. doi: 10.1016/j.enpol.2008.06.029
|
| [13] |
de Vries BJM, van Vuuren DP, Hoogwijk MM (2007) Renewable energy sources: Their global potential for the first-half of the 21st century at a global level: An integrated approach. Energy Policy 35: 2590-2610. doi: 10.1016/j.enpol.2006.09.002
|
| [14] | Ladanai S, Vinterbäck J (2009) Global Potential of Sustainable Biomass for Energy. Swedish University of Agricultural Sciences, Report 013. Uppsala. 29 p. |
| [15] |
Seiffert M, Kaltschmitt M, Miranda JA (2009) The biomethane potential in Chile. Biomass Bioenergy 33: 564-572. doi: 10.1016/j.biombioe.2008.09.005
|
| [16] | Ceciliano R (2010) WBA's report shows: The global potential of sustainable bioenergy is enough to meet the world's total energy needs. Bioenergy 3: 4-5. |
| [17] | Jacobson MZ, Delucchi MA (2009) A Path to Sustainable Energy by 2030. Sci Am 301: 58-65. |
| [18] |
Jacobson MZ, Delucchi MA (2011) Providing all global energy with wind, water, and solar power, Part I: Technologies, energy resources, quantities and areas of infrastructure, and materials. Energy Policy 39: 1154-1169. doi: 10.1016/j.enpol.2010.11.040
|
| [19] |
Masini A, Manichetti E (2012) The impact of behavioural factors in the renewable energy investment decision making process: Conceptual framework and empirical findings. Energy Policy 40: 28-38. doi: 10.1016/j.enpol.2010.06.062
|
| [20] |
Hillebrand B, Buttermann HG, Behringer JM, et al. (2006) The expansion of renewable energies and employment effects in Germany. Energy Policy 34: 3484-3494. doi: 10.1016/j.enpol.2005.06.017
|
| [21] |
Lehr U, Nitsch J, Kratzat M, et al. (2008) Renewable energy and employment in Germany. Energy Policy 36: 108-117. doi: 10.1016/j.enpol.2007.09.004
|
| [22] |
Moreno B, López AJ (2008) The effect of renewable energy on employment. The case of Asturias (Spain). Renew Sust Energ Rev 12: 732-751. doi: 10.1016/j.rser.2006.10.011
|
| [23] |
Thornley P, Rogers J, Huang Y (2008) Quantification of employment from biomass power plants. Renew Energ 33: 1922-1927. doi: 10.1016/j.renene.2007.11.011
|
| [24] |
Blanco MI, Rodrigues G (2009) Direct employment in the wind energy sector: An EU study. Energy Policy 37: 2847-2857. doi: 10.1016/j.enpol.2009.02.049
|
| [25] |
Hoffmann D (2009) Creation of regional added value by regional bioenergy resources. Renew Sust Energ Rev 13: 2419-2429. doi: 10.1016/j.rser.2009.04.001
|
| [26] |
del Río P, Burguillo M (2009) An empirical analysis of the impact of renewable energy deployment on local sustainability. Renew Sust Energ Rev 13: 1314-1325. doi: 10.1016/j.rser.2008.08.001
|
| [27] |
Openshaw K (2010) Biomass energy: Employment generation and its contribution to poverty alleviation. Biomass Bioenergy 34: 365-378. doi: 10.1016/j.biombioe.2009.11.008
|
| [28] |
Sastresa EL, Usón AA, Bribián IZ, et al. (2010) Local impact of renewables on employment: Assessment methodology and case study. Renew Sust Energ Rev 14: 679-690. doi: 10.1016/j.rser.2009.10.017
|
| [29] |
Dalton GJ, Lewis T (2011) Metrics for measuring job creation by renewable energy technologies, using Ireland as a case study. Renew Sust Energ Rev 15: 2123-2133. doi: 10.1016/j.rser.2011.01.015
|
| [30] |
Wüstenhagen R, Wolsink M, Bürer MJ (2007) Social acceptance of renewable energy innovation: An introduction to the concept. Energy Policy 35: 2683-2691. doi: 10.1016/j.enpol.2006.12.001
|
| [31] | REN21 (2011a) Renewables 2011. Global Status Report, Renewable Energy Policy Network for the 21st Century. |
| [32] | REN21 (2011b) Renewable Energy in the International Policy Process, Renewable Energy Policy Network for the 21st Century. |
| [33] |
Lund PD (2010). Fast market penetration of energy technologies in retrospect with application to clean energy futures. Appl Energ 87: 3575-358. doi: 10.1016/j.apenergy.2010.05.024
|
| [34] |
Jefferson M (2008) Accelerating the transition to sustainable energy systems. Energy Policy 36: 4116-4125. doi: 10.1016/j.enpol.2008.06.020
|
| [35] | Lund H (2010) Renewable Energy Systems. The Choice and Modeling of 100% Renewable Solutions. New York: Academic Press, Elsevier. |
| [36] |
Unruh GC (2000) Understanding carbon lock-in. Energy Policy 28: 817-830. doi: 10.1016/S0301-4215(00)00070-7
|
| [37] |
Unruh GC (2002) Escaping carbon lock-in. Energy Policy 30: 317-325. doi: 10.1016/S0301-4215(01)00098-2
|
| [38] |
Jacobsson S, Johnsson A (2000) The diffusion of renewable energy technology: an analytical framework and key issues for research. Energy Policy 28: 625-640. doi: 10.1016/S0301-4215(00)00041-0
|
| [39] | Bergek A (2010) Levelling the playing field? The influence of national wind power planning instruments on conflicts of interests in a Swedish county. Energy Policy 38: 2357-2369. |
| [40] |
Wolsink M (2000) Wind power and the NIMBY-myth: institutional capacity and the limited significance of public support. Renew Energ 21: 49-64. doi: 10.1016/S0960-1481(99)00130-5
|
| [41] | Wolsink M (2007) Planning of renewables schemes. Deliberative and fair decision-making on landscape issues instead of reproachful accusations of non-cooperation. Energy Policy 35: 2692-2704. |
| [42] |
Wolsink M (2012) The research agenda on social acceptance of distributed generation in smart grids: Renewable as common pool resources. Renew Sust Energ Rev 16: 822-835. doi: 10.1016/j.rser.2011.09.006
|
| [43] |
Peura P, Kuittinen H, Knuckey L, et al. (2014) Implementing Sustainable Energy—Four case studies. Int J Sust Economy 6: 19-44. doi: 10.1504/IJSE.2014.058516
|
| [44] |
Goldemberg J (2007) Ethanol for a Sustainable Energy Future. Science 315: 808-810. doi: 10.1126/science.1137013
|
| [45] |
Haas R, Watson J, Eichhammer W (2008) Transitions to sustainable energy systems—Introduction to the energy policy special issue. Energy Policy 36: 4009-4011. doi: 10.1016/j.enpol.2008.06.015
|
| [46] | Blarke MB (2008) From dusk till dawn. An essay about how the climate crisis has come to define sustainable energy in the context of the Danish experiment. Aalborg University. 58 p. |
| [47] |
Moriarty P, Honnery D (2009) What energy levels can the Earth sustain? Energy Policy 37: 2469-2474. doi: 10.1016/j.enpol.2009.03.006
|
| [48] | Donald Huisingh: personal communication, discussion in June 2014. |
| [49] |
Ackermann T, Andersson G, Söder L (2001) Distributed generation: a definition. Electr Pow Syst Res 57: 195-204. doi: 10.1016/S0378-7796(01)00101-8
|
| [50] |
El-Khattam W, Salama MMA (2004) Distributed generation technologies, definitions and benefits. Electr Pow Syst Res 71: 119-128. doi: 10.1016/j.epsr.2004.01.006
|
| [51] | CIRED (1999) Dispersed generation. Preliminary report of CIRED working group WG04, June, p. 9+Appendix (p.30). |
| [52] |
Pepermans G, Driesen J, Haeseldonckx D, et al. (2005) Distributed generation: definition, benefits and issues. Energy Policy 33: 787-798. doi: 10.1016/j.enpol.2003.10.004
|
| [53] | Chambers A (2001) Distributed generation: a nontechnical guide. Tulsa: PennWell, 283. |
| [54] |
Dondi P, Bayoumi D, Christoph HC, et al. (2002) Network integration of distributed power generation. J Power Sources 106: 1-9. doi: 10.1016/S0378-7753(01)01031-X
|
| [55] | International Energy Agency (IEA) (2002). Distributed generation in liberalised electricity markets. Paris. 128. |
| [56] | Dreisen J, Belmans R (2006) Distributed Generation: Challenges and Possible Solutions. 1-4244-0493-2/06/$20.00 ©2006 IEEE. Available from: http://ieeexplore.ieee.org/stamp/stamp.jsp?tp=&arnumber=1709099. |
| [57] | WADE (2014) What is DE? World Alliance for Decentralized Energy, 2014. Available from: http://www.localpower.org/deb_what.html. |
| [58] | DWPA (2014) Learn about distributed wind. Distributed Wind Association. Available from: http://distributedwind.org/home/learn-about-distributed-wind/. Visited 16.1.2014. |
| [59] | BTI (2014) Distributed Wind Energy, 2014. Available from: http://windenergyisrenewable.com/distributed-wind-energy/. |
| [60] | NWCC (2014) Distributed Wind Power Assessment, 2014. Available from: http://nationalwind.org/about-nwcc/prior-nwcc-activities/distributed-generation/. Visited 16.1.2014. |
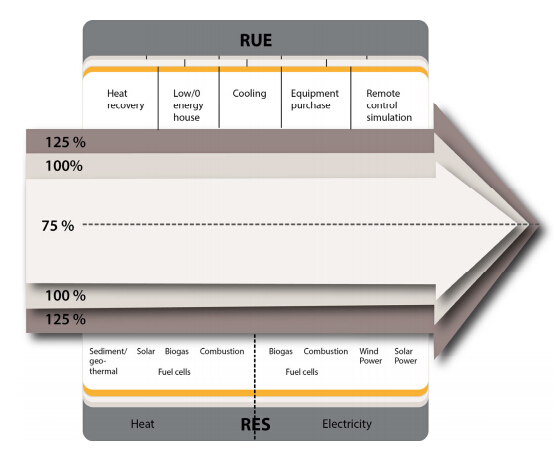
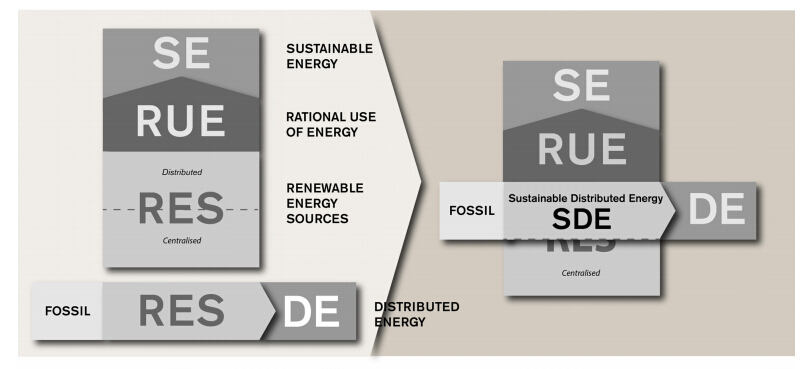
Figures(2) / Tables(1)

Pekka Peura, Patrik Sjöholm. Sustainable or Distributed Energy—or both? Clarifying the Basic Concepts of Reforming the Energy Sector[J]. AIMS Energy, 2015, 3(2): 241-254. doi: 10.3934/energy.2015.2.241







 DownLoad:
DownLoad: