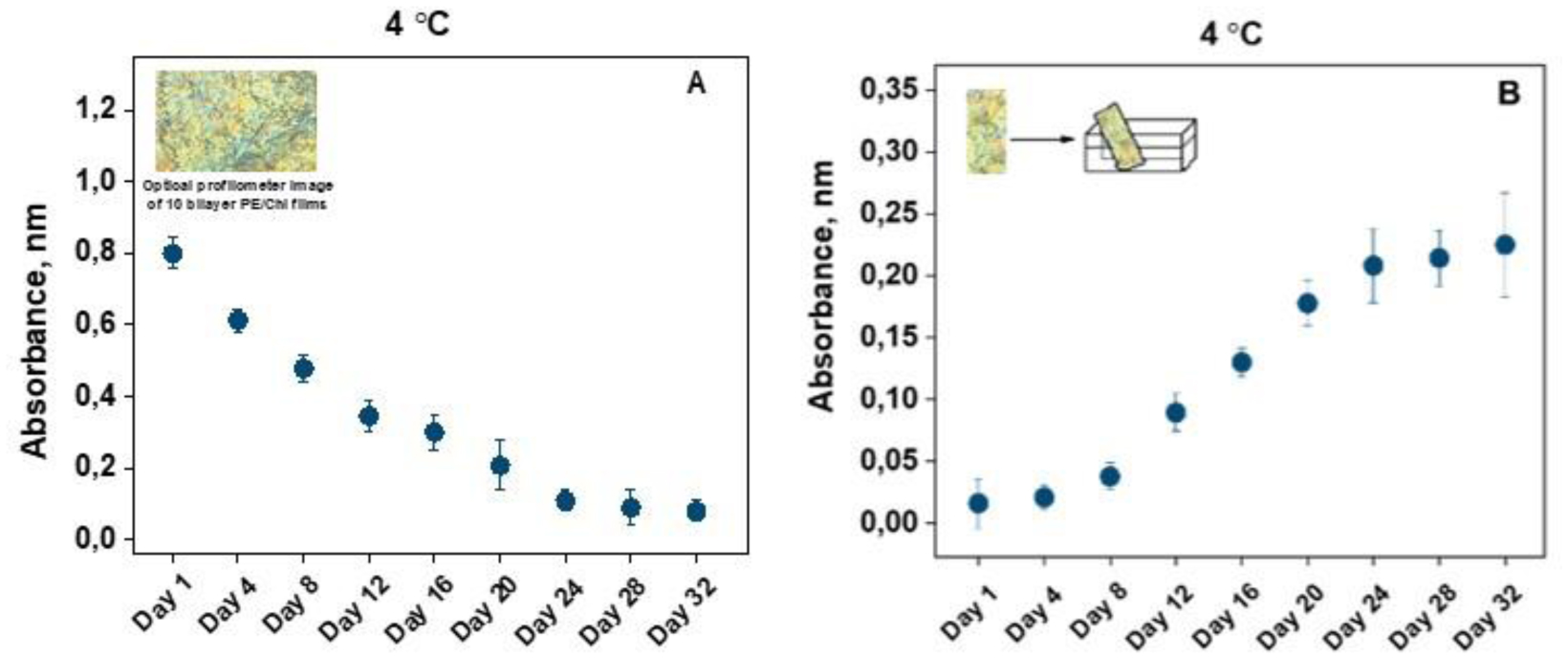
Performing their functions as transporters of oxygen and carbon dioxide in the body, human erythrocytes constantly circulate and are exposed to the constant influence of various substances, including nutrients, drugs, medical devices covered with coatings, etc. Therefore, we aim to investigate the biophysical behavior of erythrocytes obtained from healthy volunteers to observe their morphological type changes, alterations in the zeta potential, the electrical conductivity of the erythrocytes in suspensions, and hemolysis in percentages during cells senescence, both in presence and in absence of natural polyelectrolytes pectin (PE) and chitosan (Chi) in form of multilayer films (PEM-films). Being constructed using the layer–by–layer technique, films are an object of interest of many researchers because of their high potential to be incorporated in biomedicine. By applying optical profilometry, electrophoretic light scattering, and spectrophotometry, we tested the polyelectrolytes for any potential harm on the erythrocytes. Based on our results and the one-way analysis of variance (ANOVA) statistical analysis, we reached the conclusion that the above-mentioned polyelectrolytes were harmlessness; therefore, PE and Chi are suitable substances to implement in the clinical practice in the form of drug delivery carriers and medical devices coatings, thereby directly contacting with the human blood.
Citation: Nikolay Kalaydzhiev, Elena Zlatareva, Dessislava Bogdanova, Svetozar Stoichev, Avgustina Danailova. Changes in biophysical properties and behavior of aging human erythrocytes treated with natural polyelectrolytes[J]. AIMS Biophysics, 2025, 12(1): 14-28. doi: 10.3934/biophy.2025002
Performing their functions as transporters of oxygen and carbon dioxide in the body, human erythrocytes constantly circulate and are exposed to the constant influence of various substances, including nutrients, drugs, medical devices covered with coatings, etc. Therefore, we aim to investigate the biophysical behavior of erythrocytes obtained from healthy volunteers to observe their morphological type changes, alterations in the zeta potential, the electrical conductivity of the erythrocytes in suspensions, and hemolysis in percentages during cells senescence, both in presence and in absence of natural polyelectrolytes pectin (PE) and chitosan (Chi) in form of multilayer films (PEM-films). Being constructed using the layer–by–layer technique, films are an object of interest of many researchers because of their high potential to be incorporated in biomedicine. By applying optical profilometry, electrophoretic light scattering, and spectrophotometry, we tested the polyelectrolytes for any potential harm on the erythrocytes. Based on our results and the one-way analysis of variance (ANOVA) statistical analysis, we reached the conclusion that the above-mentioned polyelectrolytes were harmlessness; therefore, PE and Chi are suitable substances to implement in the clinical practice in the form of drug delivery carriers and medical devices coatings, thereby directly contacting with the human blood.

| [1] |
Kuck L, Peart JN, Simmonds MJ (2020) Active modulation of human erythrocyte mechanics. Am J Physiol Cell Physiol 319: C250-C257. https://doi.org/10.1152/ajpcell.00210.2020 
|
| [2] |
Sakamoto W, Azegami N, Konuma T, et al. (2021) Single-cell native mass spectrometry of human erythrocytes. J Anal Chem 93: 6583-6588. https://doi.org/10.1021/acs.analchem.1c00588
|
| [3] |
Giardina B, Messana I, Scatena R, et al. (1995) The multiple functions of hemoglobin. J Crit Rev Biochem Mol Biol 30: 165-196. https://doi.org/10.3109/10409239509085142
|
| [4] |
Thiagarajan P, Parker CJ, Prchal JT (2021) How do red blood cells die?. J Front Physiol 12: 655393. https://doi.org/10.3389/fphys.2021.655393
|
| [5] |
Dinarelli S, Longo G, Dietler G, et al. (2018) Erythrocyte's aging in microgravity highlights how environmental stimuli shape metabolism and morphology. Sci Rep-UK 8: 5277. https://doi.org/10.1038/s41598-018-22870-0
|
| [6] |
Goodman SR, Hughes KMH, Kakhniashvili DG, et al. (2007) The isolation of reticulocyte-free human red blood cells. J Exp Biol Med (Maywood) 232: 1470. https://doi.org/10.3181/0706-RM-163
|
| [7] |
Pretini V, Koenen MH, Kaestner L, et al. (2019) Red blood cells: chasing interactions. Front Physiol 10: 945. https://doi.org/10.3389/fphys.2019.00945
|
| [8] |
Grebowski J, Kazmierska-Grebowska P, Cichon N, et al. (2022) The effect of fullerenol C60(OH)36 on the antioxidant defense system in erythrocytes. Int J Mol Sci 23: 119. https://doi.org/10.3390/ijms23010119
|
| [9] |
Lawrence C, Meier E (2021) Chapter 17-erythrocyte disorders. Biochemical and Molecular Basis of Pediatric Disease : 529-560. https://doi.org/10.1016/B978-0-12-817962-8.00023-8
|
| [10] |
Maheshwari N, Khan FH, Mahmood R (2019) Pentachlorophenol-induced cytotoxicity in human erythrocytes: enhanced generation of ROS and RNS, lowered antioxidant power, inhibition of glucose metabolism, and morphological changes. Environ Sci Pollut Res 13: 12985-13001. https://doi.org/10.1007/s11356-019-04736-8
|
| [11] |
Piscopo M, Notariale R, Tortora F, et al. (2020) Novel insights into mercury efects on hemoglobin and membrane proteins in human erythrocytes. Molecules 25: 3278. https://doi.org/10.3390/molecules25143278
|
| [12] | Berzuini A, Bianco C, Migliorini AC, et al. (2021) Red blood cell morphology in patients with COVID-19-related anaemia. J Blood Transfus 19: 34-36. https://doi.org/10.2450/2020.0242-20 |
| [13] | Yamaguchi T, Ishimatu T (2020) Effects of cholesterol on membrane stability of human erythrocytes. J Biol Pharm Bull 43: 604-1608. https://doi.org/10.1248/bpb.b20-00435 |
| [14] |
Jan KM, Chien SH (1973) Role of surface electric charge in red blood cell interactions. J Gen Physiol 61: 638-654. https://doi.org/10.1085/jgp.61.5.638
|
| [15] |
Vincy A, Mazumder S, Amrita Banerjee I, et al. (2022) Recent progress in red blood cells-derived particles as novel bioinspired drug delivery systems: challenges and strategies for clinical translation. Front Chem 10: 905256. https://doi.org/10.3389/fchem.2022.905256
|
| [16] |
Tokumasu F, Ostera GR, Amaratunga Ch, et al. (2012) Modifications in erythrocyte membrane zeta potential by plasmodium falciparum Infection. Exp Parasitol 131: 245-251. https://doi.org/10.1016/j.exppara.2012.03.005
|
| [17] | Selim NS (2010) Comparative study on the effect of radiation on whole blood and isolated red blood cells. Romanian J Biophys 20: 127-136. |
| [18] |
Chen XY, Huang YX, Liu W, et al. (2007) Membrane surface charge and morphological and mechanical properties of young and old erythrocytes. J Curr Appl Phys 7: 94-96. https://doi.org/10.1016/j.cap.2006.11.024
|
| [19] |
Kuchinka J, Willems Ch, Telyshev DV, Groth T (2021) Control of blood coagulation by hemocompatible material surfaces—a review. Bioengineering 8: 215. https://doi.org/10.3390/bioengineering8120215
|
| [20] |
Doltchinkova V, Stoylov S, Angelova P (2021) Viper toxins affect membrane characteristics of human erythrocytes. Biophys Chem 270: 106532. https://doi.org/10.1016/j.bpc.2020.106532
|
| [21] |
Qian J, Chen Y, Wang Q, et al. (2021) Preparation and antimicrobial activity of pectin-chitosan embedding nisin microcapsules. Eur Polym J 157: 110676. https://doi.org/10.1016/j.eurpolymj.2021.110676
|
| [22] |
Xie Q, Zheng X, Li L, et al. (2021) Effect of curcumin addition on the properties of biodegradable pectin/chitosan films. Molecules 26: 2152. https://doi.org/10.3390/molecules26082152
|
| [23] |
Jovanović J, Ćirković J, Radojković A, et al. (2021) Chitosan and pectin-based films and coatings with active components for application in antimicrobial food packaging. Prog Org Coat 158: 106349. https://doi.org/10.1016/j.porgcoat.2021.106349
|
| [24] |
Jiang Z, Zhao S, Yang M, et al. (2022) Structurally stable sustained-release microcapsules stabilized by self-assembly of pectin-chitosan-collagen in aqueous two-phase system. J Food Hydrocolloids 125: 107413. https://doi.org/10.1016/j.foodhyd.2021.107413
|
| [25] |
Mallory A, Giannopoulos S, Lee P, et al. (2021) Covered stents for endovascular treatment of aortoiliac occlusive disease: a systematic review and meta-analysis. Vasc Endovasc Surg 55: 560-570. https://doi.org/10.1177/15385744211010381
|
| [26] | Kawagoe Y, Otuka F, Onozuka D, et al. (2022) Early vascular responses to abluminal biodegradable polymer-coated versus circumferential durable polymer-coated newer-generation drug-eluting stents in humans: a pathologic study. EuroIntervention 18: 1284-1294. https://doi.org/10.4244/EIJ-D-22-00650 |
| [27] |
McKenna CG, Vaughan TJ (2022) A computational framework examining the mechanical behaviour of bare and polymer-covered self-expanding laser-cut stents. J Cardiovasc Eng Tech 13: 466-480. https://doi.org/10.1007/s13239-021-00597-w
|
| [28] |
Hu B, Guo Y, Li H, et al. (2021) Recent advances in chitosan-based layer-by-layer biomaterials and their biomedical applications. Carbohyd Polym 271: 118427. https://doi.org/10.1016/j.carbpol.2021.118427
|
| [29] |
Arnon-Ripsb H, Poverenova E (2018) Improving food products' quality and storability by using layer by layer edible coatings. Trends Food Sci Tech 75: 81-92. https://doi.org/10.1016/j.tifs.2018.3.003
|
| [30] |
Grebowski J, Konarska A, Piotrowski P, et al. (2024) Kinetics of metallofullerenol reactions with the products of water radiolysis: implications for radiotherapeutics. ACS Appl Nano Mater 7: 539-549. https://doi.org/10.1021/acsanm.3c04747
|
| [31] |
Silva DCN, Jovino CN, Silva CAL, et al. (2012) Optical tweezers as a new biomedical tool to measure zeta potential of stored red blood cells. Plos One 7: e31778. https://doi.org/10.1371/journal.pone.0031778
|
| [32] |
Raat NJ, Verhoeven AJ, Mik EG (2005) The effect of storage time of human red cells on intestinal microcirculatory oxygenation in a rat isovolemic exchange model. Crit Care Med 33: 39-45. https://doi.org/10.1097/01.ccm.0000150655.75519.02
|
| [33] |
Chen KY, Lin TH, Yang CY, et al. (2018) Mechanics for the adhesion and aggregation of red blood cells on chitosan. J Mechanics 34: 725-732. https://doi.org/10.1017/jmech.2018.27
|
| [34] |
Kou S, Peters L, Mucalo M (2022) Chitosan: a review of molecular structure, bioactivities and interactions with the human body and microorganisms. J Carbohyd Polym 282: 119132. https://doi.org/10.1016/j.carbpol.2022.119132
|
| [35] |
Nadesh R, Narayanan D, Sreerekha PR, et al. (2013) Hematotoxicological analysis of surface-modified and unmodified chitosan nanoparticles. J Biomed Mater Res Part A 101A: 2957-2966. https://doi.org/10.1002/jbm.a.34591
|
| [36] |
Balan V, Verestiuc L (2014) Strategies to improve chitosan hemocompatibility: a review. Eur Polym J 53: 171-188. https://doi.org/10.1016/j.eurpolymj.2014.01.033
|
| [37] |
Zhou X, Zhang X, Zhou J, et al. (2017) An investigation of chitosan and its derivatives on red blood cell agglutination. RSC Adv 7: 12247-12254. https://doi.org/10.1039/C6RA27417
|
| [38] |
Wang W, Xue Ch, Mao X (2020) Chitosan: structural modification, biological activity and application. Int J Biol Macromol 164: 4532-4546. https://doi.org/10.1016/j.ijbiomac.2020.09.042
|
| [39] |
Azmana M, Mahmood S, Hilles AR, et al. (2021) A review on chitosan and chitosan-based bionanocomposites: Promising material for combatting global issues and its applications. Int J Biol Macromol 185: 832-848. https://doi.org/10.1016/j.ijbiomac.2021.07.023
|
| [40] |
Negm NA, Hefni HHH, Abd-Elaal AAA, et al. (2020) Advancement on modification of chitosan biopolymer and its potential applications. Int J Biol Macromol 152: 681-702. https://doi.org/10.1016/j.ijbiomac.2020.02.196
|
| [41] |
Erdogan E, Bajaj R, Lansky A, et al. (2022) Intravascular imaging for guiding in-stent restenosis and stent thrombosis therapy. J Am Heart Assoc 11: e026492. https://doi.org/10.1161/JAHA.122.026492
|
| [42] |
Nagaraja V, Schwarz K, Moss S, et al. (2020) Outcomes of patients who undergo percutaneous coronary intervention with covered stents for coronary perforation: a systematic review and pooled analysis of data. Catheter Cardio Inte 96: 1360-1366. https://doi.org/10.1002/ccd.28646
|



Figures(4) / Tables(3)

Nikolay Kalaydzhiev, Elena Zlatareva, Dessislava Bogdanova, Svetozar Stoichev, Avgustina Danailova. Changes in biophysical properties and behavior of aging human erythrocytes treated with natural polyelectrolytes[J]. AIMS Biophysics, 2025, 12(1): 14-28. doi: 10.3934/biophy.2025002







 DownLoad:
DownLoad: