Citation: Claudia Tanja Mierke. Physical role of nuclear and cytoskeletal confinements in cell migration mode selection and switching[J]. AIMS Biophysics, 2017, 4(4): 615-658. doi: 10.3934/biophy.2017.4.615

| [1] |
Albanese A, Tang PS, Chan WCW (2012) Size, shape, and surface chemistry on biological systems. Annu Rev Biomed Eng 14: 1–16. doi: 10.1146/annurev-bioeng-071811-150124

|
| [2] |
Egeblad M, Nakasone ES, Werb Z (2010) Tumors as organs: complex tissues that interface with the entire organism. Dev Cell 18: 884–901. doi: 10.1016/j.devcel.2010.05.012
|
| [3] |
Mierke CT (2014) The fundamental role of mechanical properties in the progression of cancer disease and inflammation. Rep Prog Phys 77: 076602. doi: 10.1088/0034-4885/77/7/076602
|
| [4] |
Piktel E, Niemirowicz K, Wątek M, et al. (2016) Recent insights in nanotechnology-based drugs and formulations designed for effective anti-cancer therapy. J Nanobiotechnol 14: 39. doi: 10.1186/s12951-016-0193-x
|
| [5] | Braun J, Tzschätzsch H, Körting C, et al. (2017) A compact 0.5 T MR elastography device and its application for studying viscoelasticity changes in biological tissues during progressive formalin fixation. Magn Reson Med. In press. |
| [6] | Sigrist RMS, Liau J, Kaffas AE, et al. (2017) Ultrasound elastography: review of techniques and clinical applications. Theranostics 7: 1301–1329. |
| [7] |
Mierke CT, Fischer T, Puder S, et al. (2017) Focal adhesion kinase activity is required for actomyosin contractility-based invasion of cells into dense 3D matrices. Sci Rep 7: 42780. doi: 10.1038/srep42780
|
| [8] | Jolly MK, Boareto M, Huang B, et al. (2015) Implications of the hybrid epithelial/mesenchymal phenotype in metastasis. Front Oncol 5: 00155. |
| [9] |
Jolly MK, Tripathi SC, Jia D, et al. (2016) Stability of the hybrid epithelial/mesenchymal phenotype. Oncotarget 7: 27067–27084. doi: 10.18632/oncotarget.8166
|
| [10] |
Jolly MK, Ware KE, Gilja S, et al. (2017) EMT and MET: necessary or permissive for metastasis? Mol Oncol 11: 755–769. doi: 10.1002/1878-0261.12083
|
| [11] |
Thiery JP (2002) Epithelial-mesenchymal transitions in tumour progression. Nat Rev Cancer 2: 442–454. doi: 10.1038/nrc822
|
| [12] |
Wolf K, Friedl P (2003) Tumour-cell invasion and migration: diversity and escape mechanisms. Nat Rev Cancer 3: 362–374. doi: 10.1038/nrc1075
|
| [13] |
Friedl P, Gilmour D (2009) Collective cell migration in morphogenesis, regeneration and cancer. Nat Rev Mol Cell Bio 10: 445–457. doi: 10.1038/nrm2720
|
| [14] |
Friedl P, Wolf K (2003) Tumour-cell invasion and migration: diversity and escape mechanisms. Nat Rev Cancer 3: 362–374. doi: 10.1038/nrc1075
|
| [15] |
Driscoll MK, Danuser G (2015) Quantifying modes of 3D cell migration. Trends Cell Biol 25: 749–759. doi: 10.1016/j.tcb.2015.09.010
|
| [16] | Abercrombie M, Heaysman JEM, Pegrum SM (1971) The locomotion of fibroblasts in culture. IV Electron microscopy of the leading lamella. Exp Cell Res 67: 359–367. |
| [17] |
Ridley AJ, Schwartz MA, Burridge K, et al. (2003) Cell migration: integrating signals from front to back. Science 302: 1704–1709. doi: 10.1126/science.1092053
|
| [18] |
Lauffenburger DA, Horwitz AF (1996) Cell migration: a physically integrated molecular process. Cell 84: 359–369. doi: 10.1016/S0092-8674(00)81280-5
|
| [19] |
Huttenlocher A, Sandborg RR, Horwitz AF (1995) Adhesion in cell migration. Curr Opin Cell Biol 7: 697–706. doi: 10.1016/0955-0674(95)80112-X
|
| [20] |
Geiger B, Spatz JP, Bershadsky AD (2009) Environmental sensing through focal adhesions. Nat Rev Mol Cell Biol 10: 21–33. doi: 10.1038/nrm2593
|
| [21] |
Wilson CA, Tsuchida MA, Allen GM, et al. (2010) Myosin II contributes to cell-scale actin network treadmilling through network disassembly. Nature 465: 373–377. doi: 10.1038/nature08994
|
| [22] |
Reymann AC, Boujemaa-Paterski R, Martiel JL, et al. (2012) Actin network architecture can determine myosin motor activity. Science 336: 1310–1314. doi: 10.1126/science.1221708
|
| [23] |
Petrie RJ, Yamada KM (2015) Fibroblasts lead the way: a unified view of three-dimensional cell motility. Trends Cell Biol 25: 666–674. doi: 10.1016/j.tcb.2015.07.013
|
| [24] |
Tozluoğlu M, Tournier AL, Jenkins RP, et al. (2013) Matrix geometry determines optimal cancer cell migration strategy and modulates response to interventions. Nat Cell Biol 15: 751–764. doi: 10.1038/ncb2775
|
| [25] |
Mierke CT, Rösel D, Fabry B, et al. (2008) Contractile forces in tumor cell migration. Eur J Cell Biol 87: 669–676. doi: 10.1016/j.ejcb.2008.01.002
|
| [26] |
Wolf K, Mazo I, Leung H, et al. (2003) Compensation mechanism in tumor cell migration: mesenchymal-amoeboid transition after blocking of pericellular proteolysis. J Cell Biol 160: 267–277. doi: 10.1083/jcb.200209006
|
| [27] |
Sahai E, Marshall CJ (2003) Differing modes of tumour cell invasion have distinct requirements for Rho/ROCK signalling and extracellular proteolysis. Nat Cell Biol 5: 711–719. doi: 10.1038/ncb1019
|
| [28] |
Sanz-Moreno V, Gadea G, Ahn J, et al. (2008) Rac activation and inactivation control plasticity of tumor cell movement. Cell 135: 510–523. doi: 10.1016/j.cell.2008.09.043
|
| [29] |
Neilson MP, Veltman DM, van Haastert PJ, et al. (2011) Chemotaxis: a feedback-based computational model robustly predicts multiple aspects of real cell behaviour. PLoS Biol 9: e1000618. doi: 10.1371/journal.pbio.1000618
|
| [30] |
Bagorda A, Parent CA (2008) Eukaryotic chemotaxis at a glance. J Cell Sci 121: 2621–2624. doi: 10.1242/jcs.018077
|
| [31] |
Rorth P (2011) Whence directionality: guidance mechanisms in solitary and collective cell migration. Dev Cell 20: 9–18. doi: 10.1016/j.devcel.2010.12.014
|
| [32] |
Mierke CT, Frey B, Fellner M, et al. (2011) Integrin α5β1 facilitates cancer cell invasion through enhanced contractile forces. J Cell Sci 124: 369–383. doi: 10.1242/jcs.071985
|
| [33] | Huttenlocher A, Horwitz AR (2011) Integrins in cell migration. CSH Perspect Biol 3: a005074. |
| [34] |
Hung WC, Chen SH, Paul CD, et al. (2013) Distinct signaling mechanisms regulate migration in unconfined versus confined spaces. J Cell Biol 202: 807–824. doi: 10.1083/jcb.201302132
|
| [35] |
Hung WC, Yang JR, Yankaskas CL, et al. (2016) Confinement sensing and signal optimization via Piezo1/PKA and Myosin II pathways. Cell Rep 15: 1430–1441. doi: 10.1016/j.celrep.2016.04.035
|
| [36] |
Liu YJ, Le BM, Lautenschlaeger F, et al. (2015) Confinement and low adhesion induce fast amoeboid migration of slow mesenchymal cells. Cell 160: 659–672. doi: 10.1016/j.cell.2015.01.007
|
| [37] |
Beadle C, Assanah MC, Monzo P, et al. (2008) The role of myosin II in glioma invasion of the brain. Mol Biol Cell 19: 3357–3368. doi: 10.1091/mbc.E08-03-0319
|
| [38] |
Jacobelli J, Friedman RS, Conti MA, et al. (2010) Confinement-optimized three-dimensional T cell amoeboid motility is modulated via myosin IIA-regulated adhesions. Nat Immunol 11: 953–961. doi: 10.1038/ni.1936
|
| [39] |
Han J, Rose DM, Woodside DG, et al. (2003) Integrin alpha 4 beta 1-dependent T cell migration requires both phosphorylation of the alpha 4 cytoplasmic domain to regulate the reversible binding to paxillin. J Biol Chem 278: 34845–34853. doi: 10.1074/jbc.M304691200
|
| [40] |
Charras G, Sahai E (2014) Physical influences of the extracellular environment on cell migration. Nat Rev Mol Cell Biol 15: 813–824. doi: 10.1038/nrm3897
|
| [41] |
Bergert M, Erzberger A, Desai RA, et al. (2015) Force transmission during adhesion-independent migration. Nat Cell Biol 17: 524–529. doi: 10.1038/ncb3134
|
| [42] |
Raman PS, Paul CD, Stroka KM (2013) Probing cll traction forces in confined microenvironments. Lab Chip 13: 4599–4607. doi: 10.1039/c3lc50802a
|
| [43] |
Wolenski RS (1995) Regulation of calmodulin-binding myosins. Trends Cell Biol 5: 310–316. doi: 10.1016/S0962-8924(00)89053-4
|
| [44] |
Somlyo AP, Somlyo AV (2003) Ca2+ sensitivity of smooth muscle and nonmuscle myosin II: Modulate by G proteins, kinases, and myosin phosphate. Physiol Rev 83: 1325–1358. doi: 10.1152/physrev.00023.2003
|
| [45] | Abercrombie M, Heaysman JEM, Pegrum SM (1970) The locomotion of fibroblasts in culture: I. Movements of the leading edge. Exp Cell Res 59: 393–398. |
| [46] | Bard JB, Hay ED (1975) The behavior of fibroblasts from the developing avian cornea. Morphology and movement in situ and in vitro. J Cell Biol 67: 400–418. |
| [47] |
Weiger MC, Wang CC, Krajcovic M, et al. (2009) Spontaneous phosphoinositide 3-kinase signaling dynamics drive spreading and random migration of fibroblasts. J Cell Sci 122: 313–323. doi: 10.1242/jcs.037564
|
| [48] |
Kraynov VS, Chamberlain C, Bokoch GM, et al. (2000) Localized Rac activation dynamics visualized in living cells. Science 290: 333–337. doi: 10.1126/science.290.5490.333
|
| [49] |
Nalbant P, Hodgson L, Kraynov V, et al. (2004) Activation of endogenous Cdc42 visualized in living cells. Science 305: 1615–1619. doi: 10.1126/science.1100367
|
| [50] |
Svitkina TM, Borisy GG (1999) Arp2/3 complex and actin depolymerizing factor/cofilin in dendritic organization and treadmilling of actin filament array in lamellipodia. J Cell Biol 145: 1009–1026. doi: 10.1083/jcb.145.5.1009
|
| [51] |
Small JV (1981) Organization of actin in the leading edge of cultured cells: influence of osmium tetroxide and dehydration on the ultrastructure of actin meshworks. J Cell Biol 91: 695–705. doi: 10.1083/jcb.91.3.695
|
| [52] |
Zaidel-Bar R, Ballestrem C, Kam Z, et al. (2003) Early molecular events in the assembly of matrix adhesions at the leading edge of migrating cells. J Cell Sci 116: 4605–4613. doi: 10.1242/jcs.00792
|
| [53] |
Chrzanowska-Wodnicka M, Burridge K (1996) Rho-stimulated contractility drives the formation of stress fibers and focal adhesions. J Cell Biol 133: 1403–1415. doi: 10.1083/jcb.133.6.1403
|
| [54] |
Gupton SL, Eisenmann K, Alberts AS, et al. (2007) mDia2 regulates actin and focal adhesion dynamics and organization in the lamella for efficient epithelial cell migration. J Cell Sci 120: 3475–3487. doi: 10.1242/jcs.006049
|
| [55] |
Chen WT (1981) Mechanism of retraction of the trailing edge during fibroblast movement. J Cell Biol 90: 187–200. doi: 10.1083/jcb.90.1.187
|
| [56] |
Vicente-Manzanares M, Koach MA, Whitmore L, et al. (2008) Segregation and activation of myosin IIB creates a rear in migrating cells. J Cell Biol 183: 543–554. doi: 10.1083/jcb.200806030
|
| [57] |
Bergmann JE, Kupfer A, Singer SJ (1983) Membrane insertion at the leading-edge of motile fibroblasts. Proc Natl Acad Sci USA 80: 1367–1371. doi: 10.1073/pnas.80.5.1367
|
| [58] |
Osmani N, Peglion F, Chavrier P, et al. (2010) Cdc42 localization and cell polarity depend on membrane traffic. J Cell Biol 191: 1261–1269. doi: 10.1083/jcb.201003091
|
| [59] |
Gomes ER, Jani S, Gundersen GG (2005) Nuclear movement regulated by Cdc42, MRCK, myosin, and actin flow establishes MTOC polarization in migrating cells. Cell 121: 451–463. doi: 10.1016/j.cell.2005.02.022
|
| [60] |
Petrie RJ, Yamada KM (2012) At the leading edge of three-dimensional cell migration. J Cell Sci 125: 5917–5926. doi: 10.1242/jcs.093732
|
| [61] |
Petrie RJ, Gavara N, Chadwick RS, et al. (2012) Nonpolarized signaling reveals two distinct modes of 3D cell migration. J Cell Biol 197: 439–455. doi: 10.1083/jcb.201201124
|
| [62] | Charras G, Paluch E (2008) Blebs lead the way: how to migrate without lamellipodia. Nat Rev Mol Cell Biol 9: 730–736. |
| [63] |
Lämmermann T, Bader BL, Monkley SJ, et al. (2008) Rapid leukocyte migration by integrin-independent flowing and squeezing. Nature 453: 51–55. doi: 10.1038/nature06887
|
| [64] |
Sanchez T, Chen DT, DeCamp SJ, et al. (2012) Spontaneous motion in hierarchically assembled active matter. Nature 491: 431–434. doi: 10.1038/nature11591
|
| [65] |
Ruprecht V, Wieser S, Callan-Jones A, et al. (2015) Cortical contractility triggers a stochastic switch to fast amoeboid cell motility. Cell 160: 673–685. doi: 10.1016/j.cell.2015.01.008
|
| [66] |
Davidson PM, Denais C, Bakshi MC, et al. (2014) Nuclear deformability constitutes a rate-limiting step during cell migration in 3-D environments. Cell Mol Bioeng 7: 293–306. doi: 10.1007/s12195-014-0342-y
|
| [67] |
Harada T, Swift J, Irianto J, et al. (2014) Nuclear lamin stiffness is a barrier to 3D migration, but softness can limit survival. J Cell Biol 204: 669–682. doi: 10.1083/jcb.201308029
|
| [68] |
Wolf K, Lindert MT, Krause M, et al. (2013) Physical limits of cell migration: control by ECM space and nuclear deformation and tuning by proteolysis and traction force. J Cell Biol 201: 1069–1084. doi: 10.1083/jcb.201210152
|
| [69] |
Bernards R, Weinberg RA (2002) Metastasis genes: A progression puzzle. Nature 418: 823. doi: 10.1038/418823a
|
| [70] |
Heasman SJ, Ridley AJ (2008) Mammalian Rho GTPases: new insights into their functions from in vivo studies. Nat Rev Mol Cell Biol 9: 690–701. doi: 10.1038/nrm2476
|
| [71] |
Ladwein M, Rottner K (2008) On the Rho'd: the regulation of membrane protrusions by Rho-GTPases. FEBS Lett 582: 2066–2074. doi: 10.1016/j.febslet.2008.04.033
|
| [72] |
Aspenström P, Fransson A, Saras J (2004) Rho GTPases have diverse effects on the organization of the actin filament system. Biochem J 377: 327–337. doi: 10.1042/bj20031041
|
| [73] |
Ridley AJ, Paterson HF, Johnston CL, et al. (1992) The small GTP-binding protein rac regulates growth factor-induced membrane ruffling. Cell 70: 401–410. doi: 10.1016/0092-8674(92)90164-8
|
| [74] |
Nobes CD, Hall A (1995) Rho, rac, and cdc42 GTPases regulate the assembly of multimolecular focal complexes associated with actin stress fibers, lamellipodia, and filopodia. Cell 81: 53–62. doi: 10.1016/0092-8674(95)90370-4
|
| [75] |
Monypenny J, Zicha D, Higashida C, et al. (2009) Cdc42 and Rac family GTPases regulate mode and speed but not direction of primary fibroblast migration during platelet-derived growth factor-dependent chemotaxis. Mol Cell Biol 29: 2730–2747. doi: 10.1128/MCB.01285-08
|
| [76] |
Krugmann S, Jordens I, Gevaert K, et al. (2001) Cdc42 induces filopodia by promoting the formation of an IRSp53: Mena complex. Curr Biol 11: 1645–1655. doi: 10.1016/S0960-9822(01)00506-1
|
| [77] |
Jaffe AB, Hall A (2002) Rho GTPases in transformation and metastasis. Adv Cancer Res 84: 57–80. doi: 10.1016/S0065-230X(02)84003-9
|
| [78] |
Hall A (1998) Rho GTPases and the actin cytoskeleton. Science 279: 509–514. doi: 10.1126/science.279.5350.509
|
| [79] |
Allen WE, Zicha D, Ridley AJ, et al. (1998) A role for Cdc42 in macrophage chemotaxis. J Cell Biol 141: 1147–1157. doi: 10.1083/jcb.141.5.1147
|
| [80] |
Nobes CD, Hall A (1999) Rho GTPases control polarity, protrusion, and adhesion during cell movement. J Cell Biol 144: 1235–1244. doi: 10.1083/jcb.144.6.1235
|
| [81] |
Amano M, Chihara K, Kimura K, et al. (1997) Formation of actin stress fibers and focal adhesions enhanced by Rho-kinase. Science 275: 1308–1311. doi: 10.1126/science.275.5304.1308
|
| [82] |
Watanabe N, Kato T, Fujita A, et al. (1999) Cooperation between mDia1 and ROCK in Rho-induced actin reorganization. Nat Cell Biol 1: 136–143. doi: 10.1038/11056
|
| [83] |
Cox EA, Sastry SK, Huttenlocher A (2001) Integrin-mediated adhesion regulates cell polarity and membrane protrusion through the Rho family of GTPases. Mol Biol Cell 12: 265–277. doi: 10.1091/mbc.12.2.265
|
| [84] |
Fukata M, Kaibuchi K (2001) Rho-family GTPases in Cadherin-mediated cell-cell adhesion. Nat Rev Mol Cell Bio 2: 887–897. doi: 10.1038/35103068
|
| [85] |
Doyle AD, Wang FW, Matsumoto K, et al. (2009) One-dimensional topography underlies three-dimensional fibrillar cell migration. J Cell Biol 184: 481–490. doi: 10.1083/jcb.200810041
|
| [86] |
Even-Ram S, Doyle AD, Conti MA, et al. (2007) Myosin IIA regulates cell motility and actomyosin-microtubule crosstalk. Nat Cell Biol 9: 299–309. doi: 10.1038/ncb1540
|
| [87] |
Zhong C, Chrzanowska-Wodnicka M, Brown J, et al. (1998) Rho-mediated contractility exposes a cryptic site in fibronectin and induces fibronectin matrix assembly. J Cell Biol 141: 539–551. doi: 10.1083/jcb.141.2.539
|
| [88] |
Arthur WT, Burridge K (2001) RhoA inactivation by p190RhoGAP regulates cell spreading and migration by promoting membrane protrusion and polarity. Mol Biol Cell 12: 2711–2720. doi: 10.1091/mbc.12.9.2711
|
| [89] | Chrzanowska-Wodnicka M, Burridge K (1994) Tyrosine phosphorylation is involved in reorganization of the actin cytoskeleton in response to serum or LPA stimulation. J Cell Sci 107: 3643–3654. |
| [90] |
Provenzano PP, Inman DR, Eliceiri KW, et al. (2008) Contact guidance mediated three-dimensional cell migration is regulated by Rho/ROCK-dependent matrix reorganization. Biophys J 95: 5374–5384. doi: 10.1529/biophysj.108.133116
|
| [91] |
Riching KM, Cox BL, Salick MR, et al. (2014) 3D collagen alignment limits protrusions to enhance breast cancer cell persistence. Biophys J 107: 2546–2558. doi: 10.1016/j.bpj.2014.10.035
|
| [92] |
Thomas DG, Yenepalli A, Denais CM, et al. (2015) Non-muscle myosin IIB is critical for nuclear translocation during 3D invasion. J Cell Biol 210: 583–594. doi: 10.1083/jcb.201502039
|
| [93] | Paul CD, Mistriotis P, Konstantopoulos K (2017) Cancer cell motility: lessons from migration in confined spaces. Nat Rev Cancer 17: 131–140. |
| [94] |
Lombardi ML, Jaalouk DE, Shanahan CM, et al. (2011) The interaction between nesprins and sun proteins at the nuclear envelope is critical for force transmission between the nucleus and cytoskeleton. J Biol Chem 286: 26743–26753. doi: 10.1074/jbc.M111.233700
|
| [95] |
Mierke CT, Zitterbart DP, Kollmannsberger P, et al. (2008) Breakdown of the endothelial barrier function in tumor cell transmigration. Biophys J 94: 2832–2846. doi: 10.1529/biophysj.107.113613
|
| [96] | Eddy RJ, Pierini LM, Matsumura F, et al. (2000) Ca2+-dependent myosin II activation is required for uropod retraction during neutrophil migration. J Cell Sci 113: 1287–1298. |
| [97] |
Poincloux R, Collin O, Lizárraga F, et al. (2011) Contractility of the cell rear drives invasion of breast tumor cells in 3D Matrigel. Proc Natl Acad Sci USA 108: 1943–1948. doi: 10.1073/pnas.1010396108
|
| [98] |
Olins AL, Hoang TV, Zwerger M, et al. (2009) The LINC-less granulocyte nucleus. Eur J Cell Biol 88: 203–214. doi: 10.1016/j.ejcb.2008.10.001
|
| [99] |
Machacek M, Hodgson L, Welch C, et al. (2009) Coordination of Rho GTPase activities during cell protrusion. Nature 461: 99–103. doi: 10.1038/nature08242
|
| [100] |
Das T, Safferling K, Rausch S, et al. (2015) A molecular mechanotransduction pathway regulates collective migration of epithelial cells. Nat Cell Biol 17: 276–287. doi: 10.1038/ncb3115
|
| [101] |
Yi CL, Troutman S, Fera D, et al. (2011) A tight junction-associated merlin-angiomotin complex mediates Merlin's regulation of mitogenic signaling and tumor suppressive functions. Cancer Cell 19: 527–540. doi: 10.1016/j.ccr.2011.02.017
|
| [102] |
Ramel D, Wang XB, Laflamme C, et al. (2013) Rab11 regulates cell-cell communication during collective cell movements. Nat Cell Biol 15: 317–324. doi: 10.1038/ncb2681
|
| [103] |
Gaggioli C, Hooper S, Hidalgo-Carcedo C, et al. (2007) Fibroblast-led collective invasion of carcinoma cells with differing roles for RhoGTPases in leading and following cells. Nat Cell Biol 9: 1392–1400. doi: 10.1038/ncb1658
|
| [104] |
Wang XB, He L, Wu YI, et al. (2010) Light-mediated activation reveals a key role for Rac in collective guidance of cell movement in vivo. Nat Cell Biol 12: 591–597. doi: 10.1038/ncb2061
|
| [105] |
Hidalgo-Carcedo C, Hooper S, Chaudhry SI, et al. (2011) Collective cell migration requires suppression of actomyosin at cell-cell contacts mediated by DDR1 and the cell polarity regulators Par3 and Par6. Nat Cell Biol 13: 49–58. doi: 10.1038/ncb2133
|
| [106] |
Ng MR, Besser A, Danuser G, et al. (2012) Substrate stiffness regulates cadherin-dependent colle cadherin-dependent collective migration through myosin-II contractility. J Cell Biol 199: 545–563. doi: 10.1083/jcb.201207148
|
| [107] |
Reffay M, Parrini MC, Cochet-Escartin O, et al. (2014) Interplay of RhoA and mechanical forces in collective cell migration driven by leader cells. Nat Cell Biol 16: 217–223. doi: 10.1038/ncb2917
|
| [108] |
Peglion F, Llense F, Etienne-Manneville S (2014) Adherens junction treadmilling during collective migration. Nat Cell Biol 16: 639–651. doi: 10.1038/ncb2985
|
| [109] |
Raupach C, Zitterbart DP, Mierke CT, et al. (2007) Stress fluctuations and motion of cytoskeletal-bound markers. Phys Rev E 76: 011918. doi: 10.1103/PhysRevE.76.011918
|
| [110] |
Tambe DT, Hardin CC, Angelini TE, et al. (2011) Collective cell guidance by cooperative intercellular forces. Nat Mater 10: 469–475. doi: 10.1038/nmat3025
|
| [111] |
Trepat X, Wasserman MR, Angelini TE, et al. (2009) Physical forces during collective cell migration. Nat Phys 5: 426–430. doi: 10.1038/nphys1269
|
| [112] |
Jaffe AB, Hall A (2005) Rho GTPases: biochemistry and biology. Annu Rev Cell Dev Biol 21: 247–269. doi: 10.1146/annurev.cellbio.21.020604.150721
|
| [113] |
Schmidt A, Hall A (2002) Guanine nucleotide exchange factors for Rho GTPases: turning on the switch. Gene Dev 16: 1587–1609. doi: 10.1101/gad.1003302
|
| [114] |
Otani T, Ichii T, Aono S, et al. (2006) Cdc42 GEF Tuba regulates the junctional configuration of simple epithelial cells. J Cell Biol 175: 135–146. doi: 10.1083/jcb.200605012
|
| [115] |
Harris KP, Tepass U (2010) Cdc42 and vesicle trafficking in polarized cells. Traffic 11: 1272–1279. doi: 10.1111/j.1600-0854.2010.01102.x
|
| [116] |
Citi S, Spadaro D, Schneider Y, et al. (2011) Regulation of small GTPases at epithelial cell-cell junctions. Mol Membr Biol 28: 427–444. doi: 10.3109/09687688.2011.603101
|
| [117] |
Rohatgi R, Ho HY, Kirschner MW (2000) Mechanism of N-WASP activation by CDC42 and phosphatidylinositol 4, 5-bisphosphate. J Cell Biol 150: 117–120. doi: 10.1083/jcb.150.6.F117
|
| [118] |
Hall A (2005) Rho GTPases and the control of cell behaviour. Biochem Soc Trans 33: 891–895. doi: 10.1042/BST0330891
|
| [119] |
Aspenström P, Richnau N, Johansson AS (2006) The diaphanous-related formin DAAM1 collaborates with the Rho GTPases RhoA and Cdc42, CIP4 and Src in regulating cell morphogenesis and actin dynamics. Exp Cell Res 312: 2180–2194. doi: 10.1016/j.yexcr.2006.03.013
|
| [120] |
Aspenstrom P (1997) A Cdc42 target protein with homology to the non-kinase domain of FER has a potential role in regulating the actin cytoskeleton. Curr Biol 7: 479–487. doi: 10.1016/S0960-9822(06)00219-3
|
| [121] |
Tian L, Nelson DL, Stewart DM (2000) Cdc42-interacting protein 4 mediates binding of the Wiskott-Aldrich syndrome protein to microtubules. J Biol Chem 275: 7854–7861. doi: 10.1074/jbc.275.11.7854
|
| [122] |
Yanai M, Kenyon CM, Butler JP, et al. (1996) Intracellular pressure is a motive force for cell motion in Amoeba proteus. Cell Motil Cytoskel 33: 22–29. doi: 10.1002/(SICI)1097-0169(1996)33:1<22::AID-CM3>3.0.CO;2-K
|
| [123] | Petrie RJ, Koo H (2014) Direct measurement of intracellular pressure. Curr Protoc Cell Biol 63: 12–19. |
| [124] |
Petrie RJ, Koo H, Yamada KM (2014) Generation of compartmentalized pressure by a nuclear piston governs cell motility in a 3D matrix. Science 345: 1062–1065. doi: 10.1126/science.1256965
|
| [125] |
Stroka KM, Jiang H, Chen SH, et al. (2014) Water permeation drives tumor cell migration in confined microenvironments. Cell 157: 611–623. doi: 10.1016/j.cell.2014.02.052
|
| [126] |
Chartier L, Rankin LL, Allen RE, et al. (1991) Calyculin-A increases the level of protein phosphorylation and changes the shape of 3T3 fibroblasts. Cell Motil Cytoskel 18: 26–40. doi: 10.1002/cm.970180104
|
| [127] |
Goldman R, Goldman A, Green K, et al. (1985) Intermediate filaments: possible functions as cytoskeletal connecting links between the nucleus and the cell surface. Ann NY Acad Sci 455: 1–17. doi: 10.1111/j.1749-6632.1985.tb50400.x
|
| [128] |
Lehto VP, Virtanen I, Kurki P (1978) Intermediate filaments anchor the nuclei in nuclear monolayers of cultured human fibroblasts. Nature 272: 175–177. doi: 10.1038/272175a0
|
| [129] |
Janmey PA, Euteneuer U, Traub P, et al. (1991) Viscoelastic properties of vimentin compared with other filamentous biopolymer networks. J Cell Biol 113: 155–160. doi: 10.1083/jcb.113.1.155
|
| [130] |
Deguchi S, Ohashi T, Sato M (2006) Tensile properties of single stress fibers isolated from cultured vascular smooth muscle cells. J Biomech 39: 2603–2610. doi: 10.1016/j.jbiomech.2005.08.026
|
| [131] |
Dupin I, Sakamoto Y, Etienne-Manneville S (2011) Cytoplasmic intermediate filaments mediate actin-driven positioning of the nucleus. J Cell Sci 124: 865–872. doi: 10.1242/jcs.076356
|
| [132] |
Petrie RJ, Harlin HM, Korsak LIT, et al. (2017) Activating the nuclear piston mechanism of 3D migration in tumor cells. J Cell Biol 216: 93–100. doi: 10.1083/jcb.201605097
|
| [133] | Sung BH, Ketova T, Hoshino D, et al. (2004) Directional cell movement through tissues is controlled by exosome secretion. Nat Commun 6: 7164. |
| [134] |
Gundersen GG, Bulinski JC (1988) Selective stabilization of microtubules oriented toward the direction of cell migration. Proc Natl Acad Sci USA 85: 5946–5950. doi: 10.1073/pnas.85.16.5946
|
| [135] |
Kupfer A, Louvard D, Singer SJ (1982) Polarization of the Golgi apparatus and the microtubule-organizing center in cultured fibroblasts at the edge of an experimental wound. Proc Natl Acad Sci USA 79: 2603–2607. doi: 10.1073/pnas.79.8.2603
|
| [136] | Vasiliev JM, Gelfand IM, Domnina LV, et al. (1970) Effect of colcemid on the locomotory behaviour of fibroblasts. J Embryol Exp Morphol 24: 625–640. |
| [137] | Wu C, Asokan SB, Berginski ME, et al. (2012) Arp2/3 is critical for lamellipodia and response to extracellular matrix cues but is dispensable for chemotaxis. Cell 148: 973–987. |
| [138] |
Wu YI, Frey D, Lungu OI, et al. (2009) A genetically encoded photoactivatable Rac controls the motility of living cells. Nature 461: 104–108. doi: 10.1038/nature08241
|
| [139] |
Welf ES, Ahmed S, Johnson HE, et al. (2012) Migrating fibroblasts reorient directionality by a metastable, PI3K-dependent mechanism. J Cell Biol 197: 105–114. doi: 10.1083/jcb.201108152
|
| [140] |
Giannone G, Dubin-Thaler BJ, Döbereiner HG, et al. (2004) Periodic lamellipodial contractions correlate with rearward actin waves. Cell 116: 431–443. doi: 10.1016/S0092-8674(04)00058-3
|
| [141] |
Ponti A, Machacek M, Gupton SL, et al. (2004) Two distinct actin networks drive the protrusion of migrating cells. Science 305: 1782–1786. doi: 10.1126/science.1100533
|
| [142] |
Friedl P, Wolf K (2009) Proteolytic interstitial cell migration: a five-step process. Cancer Metastasis Rev 28: 129–135. doi: 10.1007/s10555-008-9174-3
|
| [143] |
Danen EH, van Rheenen J, Franken W, et al. (2005) Integrins control motile strategy through a Rho-cofilin pathway. J Cell Biol 169: 515–526. doi: 10.1083/jcb.200412081
|
| [144] |
Doyle AD, Kutys ML, Conti MA, et al. (2012) Micro-environmental control of cell migration-myosin IIA is required for efficient migration in fibrillar environments through control of cell adhesion dynamics. J Cell Sci 125: 2244–2256. doi: 10.1242/jcs.098806
|
| [145] |
Lämmermann T, Sixt M (2009) Mechanical modes of 'amoeboid' cell migration. Curr Opin Cell Biol 21: 636–644. doi: 10.1016/j.ceb.2009.05.003
|
| [146] |
Renkawitz J, Schumann K, Weber M, et al. (2009) Adaptive force transmission in amoeboid cell migration. Nat Cell Biol 11: 1438–1443. doi: 10.1038/ncb1992
|
| [147] |
Charras GT, Yarrow JC, Horton MA, et al. (2005) Non-equilibration of hydrostatic pressure in blebbing cells. Nature 435: 365–369. doi: 10.1038/nature03550
|
| [148] |
Bergert M, Chandradoss SD, Desai RA, et al. (2012) Cell mechanics control rapid transitions between blebs and lamellipodia during migration. Proc Natl Acad Sci USA 109: 14434–14439. doi: 10.1073/pnas.1207968109
|
| [149] |
Petrie RJ, Yamada KM (2016) Multiple mechanisms of 3D migration: The origins of plasticity. Curr Opin Cell Biol 42: 7–12. doi: 10.1016/j.ceb.2016.03.025
|
| [150] |
Doyle AD, Carvajal N, Jin A, et al. (2015) Local 3D matrix microenvironment regulates cell migration through spatiotemporal dynamics of contractility-dependent adhesions. Nat Commun 6: 8720. doi: 10.1038/ncomms9720
|
| [151] |
Ray U, Roy SS, Chowdhury SR (2017) Lysophosphatidic acid promotes epithelial to mesenchymal transition in ovarian cancer cells by repressing SIRT1. Cell Physiol Biochem 41: 795–805. doi: 10.1159/000458744
|
| [152] |
Vincent T, Neve EP, Johnson JR, et al. (2009) A SNAIL1-SMAD3/4 transcriptional repressor complex promotes TGF-beta mediated epithelial-mesenchymal transition. Nat Cell Biol 11: 943–950. doi: 10.1038/ncb1905
|
| [153] |
Huang YL, Tung CK, Zheng A, et al. (2015) Interstitial flows promote amoeboid over mesenchymal motility of breast cancer cells revealed by a three dimensional microfluidic model. Integr Biol 7: 1402–1411. doi: 10.1039/C5IB00115C
|
| [154] |
Laurent VM, Fodil R, Cañadas P, et al. (2003) Partitioning of cortical and deep cytoskeleton responses from transient magnetic bead twisting. Ann Biomed Eng 31: 1263–1278. doi: 10.1114/1.1616932
|
| [155] |
Laevsky G, Knecht DA (2003) Cross-linking of actin filaments by myosin II is a major contributor to cortical integrity and cell motility in restrictive environments. J Cell Sci 116: 3761–3770. doi: 10.1242/jcs.00684
|
| [156] |
Small JV, Rottner K, Kaverina I, et al. (1998) Assembling an actin cytoskeleton for cell attachment and movement. Biochim Biophys Acta 1404: 271–281. doi: 10.1016/S0167-4889(98)00080-9
|
| [157] |
Tojkander S, Gateva G, Lappalainen P (2012) Actin stress fibers-assembly, dynamics and biological roles. J Cell Sci 125: 1855–1864. doi: 10.1242/jcs.098087
|
| [158] |
Vallenius T (2013) Actin stress fibre subtypes in mesenchymal-migrating cells. Open Biol 3: 130001. doi: 10.1098/rsob.130001
|
| [159] |
Kassianidou E, Kumar S (2015) A biomechanical perspective on stress fiber structure and function. Biochim Biophys Acta 1853: 3065–3074. doi: 10.1016/j.bbamcr.2015.04.006
|
| [160] |
Klimova Z, Braborec V, Maninova M, et al. (2016) Symmetry breaking in spreading RAT2 fibroblasts requires the MAPK/ERK pathway scaffold RACK1 that integrates FAK, p190A- RhoGAP and ERK2 signaling. Biochim Biophys Acta 1863: 2189–2200. doi: 10.1016/j.bbamcr.2016.05.013
|
| [161] |
Vicente-Manzanares M, Newell-Litwa K, Bachir AI, et al. (2011) Myosin IIA/IIB restrict adhesive and protrusive signaling to generate front-back polarity in migrating cells. J Cell Biol 193: 381–396. doi: 10.1083/jcb.201012159
|
| [162] |
Prager-Khoutorsky M, Lichtenstein A, Krishnan R, et al. (2011) Fibroblast polarization is a matrix-rigidity-dependent process controlled by focal adhesion mechanosensing. Nat Cell Biol 13: 1457–1465. doi: 10.1038/ncb2370
|
| [163] |
Iwanicki MP, Vomastek T, Tilghman RW, et al. (2008) FAK, PDZ-RhoGEF and ROCKII cooperate to regulate adhesion movement and trailing-edge retraction in fibroblasts. J Cell Sci 121: 895–905. doi: 10.1242/jcs.020941
|
| [164] |
Kumar S, Maxwell IZ, Heisterkamp A, et al. (2006) Viscoelastic retraction of single living stress fibers and its impact on cell shape, cytoskeletal organization, and extracellular matrix mechanics. Biophys J 90: 3762– 3773. doi: 10.1529/biophysj.105.071506
|
| [165] |
Pellegrin S, Mellor H (2007) Actin stress fibres. J Cell Sci 120: 3491–3499. doi: 10.1242/jcs.018473
|
| [166] | Mierke CT, Sauer F, Grosser S, et al. (2017) The two faces of enhanced stroma: Stroma acts as a tumor promoter and a steric obstacle. NMR Biomed. In press. |
| [167] |
Fischer T, Wilharm N, Hayn A, et al. (2017) Matrix and cellular mechanical properties are the driving factors for facilitating human cancer cell motility into 3D engineered matrices. Converg Sci Phys Oncol 3: 044003. doi: 10.1088/2057-1739/aa8bbb
|
| [168] |
Oakes PW, Beckham Y, Stricker J, et al. (2012) Tension is required but not sufficient for focal adhesion maturation without a stress fiber template. J Cell Biol 196: 363–374. doi: 10.1083/jcb.201107042
|
| [169] |
Kovac B, Teo JL, Makela TP, et al. (2013) Assembly of non-contractile dorsal stress fibers requires alpha-actinin-1 and Rac1 in migrating and spreading cells. J Cell Sci 126: 263–273. doi: 10.1242/jcs.115063
|
| [170] |
Feng Y, Ngu H, Alford SK, et al. (2013) Alpha-actinin1 and 4 tyrosine phosphorylation is critical for stress fiber establishment, maintenance and focal adhesion maturation. Exp Cell Res 319: 1124–1135. doi: 10.1016/j.yexcr.2013.02.009
|
| [171] |
Hotulainen P, Lappalainen P (2006) Stress fibers are generated by two distinct actin assembly mechanisms in motile cells. J Cell Biol 173: 383–394. doi: 10.1083/jcb.200511093
|
| [172] |
Burnette DT, Shao L, Ott C, et al. (2014) A contractile and counterbalancing adhesion system controls the 3D shape of crawling cells. J Cell Biol 205: 83–96. doi: 10.1083/jcb.201311104
|
| [173] |
Tojkander S, Gateva G, Schevzov G, et al. (2011) A molecular pathway for myosin II recruitment to stress fibers. Curr Biol 21: 539–550. doi: 10.1016/j.cub.2011.03.007
|
| [174] |
Gunning PW, Hardeman EC, Lappalainen P, et al. (2015) Tropomyosin-master regulator of actin filament function in the cytoskeleton. J Cell Sci 128: 2965–2974. doi: 10.1242/jcs.172502
|
| [175] |
Blanchoin L, Boujemaa-Paterski R, Sykes C, et al. (2014) Actin dynamics, architecture, and mechanics in cell motility. Physiol Rev 94: 235–263. doi: 10.1152/physrev.00018.2013
|
| [176] |
Khatau SB, Hale CM, Stewart-Hutchinson PJ, et al. (2009) A perinuclear actin cap regulates nuclear shape. Proc Natl Acad Sci USA 106: 19017–19022. doi: 10.1073/pnas.0908686106
|
| [177] |
Krause M, te Riet J, Wolf K (2013) Probing the compressibility of tumor cell nuclei by combined atomic force-confocal microscopy. Phys Biol 10: 065002. doi: 10.1088/1478-3975/10/6/065002
|
| [178] |
Thiam HR, Vargas P, Carpi N, et al. (2016) Perinuclear Arp2/3-driven actin polymerization enables nuclear deformation to facilitate cell migration through complex environments. Nat Commun 7: 10997. doi: 10.1038/ncomms10997
|
| [179] |
Kim DH, Cho S, Wirtz D (2014) Tight coupling between nucleus and cell migration through the perinuclear actin cap. J Cell Sci 127: 2528–2541. doi: 10.1242/jcs.144345
|
| [180] |
Gundersen GG, Worman HJ (2013) Nuclear positioning. Cell 152: 1376–1389. doi: 10.1016/j.cell.2013.02.031
|
| [181] |
Maninova M, Vomastek T (2016) Dorsal stress fibers, transverse actin arcs, and perinuclear actin fibers form an interconnected network that induces nuclear movement in polarizing fibroblasts. FEBS J 283: 3676–3693. doi: 10.1111/febs.13836
|
| [182] |
Luxton GWG, Gundersen GG (2011) Orientation and function of the nuclear-centrosomal axis during cell migration. Curr Opin Cell Biol 23: 579–588. doi: 10.1016/j.ceb.2011.08.001
|
| [183] |
Dupin I, Etienne-Manneville S (2011) Nuclear positioning: mechanisms and functions. Int J Biochem Cell Biol 43: 1698–707. doi: 10.1016/j.biocel.2011.09.004
|
| [184] |
Isermann P, Lammerding J (2013) Nuclear mechanics and mechanotransduction in health and disease. Curr Biol 23: R1113–R1121. doi: 10.1016/j.cub.2013.11.009
|
| [185] |
Luxton GW, Gomes ER, Folker ES, et al. (2010) Linear arrays of nuclear envelope proteins harness retrograde actin flow for nuclear movement. Science 329: 956–959. doi: 10.1126/science.1189072
|
| [186] | Steinbock FA, Wiche G (1999) Plectin: a cytolinker by design. Biol Chem 380: 151–158. |
| [187] |
Mierke CT, Kollmannsberger P, Zitterbart DP, et al. (2008) Mechano-coupling and regulation of contractility by the vinculin tail domain. Biophys J 94: 661–670. doi: 10.1529/biophysj.107.108472
|
| [188] |
Mierke CT (2013) Phagocytized beads reduce the α5β1 integrin facilitated invasiveness of cancer cells by regulating cellular stiffness. Cell Biochem Biophys 66: 599–622. doi: 10.1007/s12013-012-9506-3
|
| [189] |
Maniotis AJ, Chen CS, Ingber DE (1997) Demonstration of mechanical connections between integrins, cytoskeletal filaments, and nucleoplasm that stabilize nuclear structure. Proc Natl Acad Sci USA 94: 849–854. doi: 10.1073/pnas.94.3.849
|
| [190] |
Brosig M, Ferralli J, Gelman L, et al. (2010) Interfering with the connection between the nucleus and the cytoskeleton affects nuclear rotation, mechanotransduction and myogenesis. Int J Biochem Cell Biol 42: 1717–1728. doi: 10.1016/j.biocel.2010.07.001
|
| [191] |
Levy JR, Holzbaur ELF (2008) Dynein drives nuclear rotation during forward progression of motile fibroblasts. J Cell Sci 121: 3187–3195. doi: 10.1242/jcs.033878
|
| [192] |
Vicente-Manzanares M, Zareno J, Whitmore L, et al. (2007) Regulation of protrusion, adhesion dynamics, and polarity by myosins IIA and IIB in migrating cells. J Cell Biol 176: 573–580. doi: 10.1083/jcb.200612043
|
| [193] |
Kim DH, Chambliss AB, Wirtz D (2013) The multi-faceted role of the actin cap in cellular mechanosensation and mechanotransduction. Soft Matter 9: 5516–5523. doi: 10.1039/c3sm50798j
|
| [194] |
Kim DH, Khatau SB, Feng YF, et al. (2012) Actin cap associated focal adhesions and their distinct role in cellular mechanosensing. Sci Rep 2: 555. doi: 10.1038/srep00555
|
| [195] |
Chaffer CL, Weinberg RA (2011) A perspective on cancer cell metastasis. Science 331: 1559–1564. doi: 10.1126/science.1203543
|
| [196] |
Gupta GP, Massague J (2006) Cancer metastasis: building a framework. Cell 127: 679–695. doi: 10.1016/j.cell.2006.11.001
|
| [197] |
Thiery JP, Acloque H, Huang RY, et al. (2009) Epithelial-mesenchymal transitions in development and disease. Cell 139: 871–890. doi: 10.1016/j.cell.2009.11.007
|
| [198] |
Wirtz D, Konstantopoulos K, Searson PC (2011) The physics of cancer: the role of physical interactions and mechanical forces in metastasis. Nat Rev Cancer 11: 512–522. doi: 10.1038/nrc3080
|
| [199] |
Starr DA, Fridolfsson HN (2010) Interactions between nuclei and the cytoskeleton are mediated by SUN-KASH nuclear-envelope bridges. Annu Rev Cell Dev Biol 26: 421–444. doi: 10.1146/annurev-cellbio-100109-104037
|
| [200] |
Nagayama K, Yamazaki S, Yahiro Y, et al. (2014) Estimation of the mechanical connection between apical stress fibers and the nucleus in vascular smooth muscle cells cultured on a substrate. J Biomech 47: 1422–1429. doi: 10.1016/j.jbiomech.2014.01.042
|
| [201] |
Vishavkarma R, Raghavan S, Kuyyamudi C, et al. (2014) Role of actin filaments in correlating nuclear shape and cell spreading. PLoS One 9: e107895. doi: 10.1371/journal.pone.0107895
|
| [202] |
Chambliss AB, Khatau SB, Erdenberger N, et al. (2013) The LINC-anchored actin cap connects the extracellular milieu to the nucleus for ultrafast mechanotransduction. Sci Rep 3: 1087. doi: 10.1038/srep01087
|
| [203] |
Lin HH, Lin HK, Lin IH, et al. (2015) Mechanical phenotype of cancer cells: cell softening and loss of stiffness sensing. Oncotarget 6: 20946–20958. doi: 10.18632/oncotarget.4173
|
| [204] |
Tamiello C, Bouten CV, Baaijens FP (2015) Competition between cap and basal actin fiber orientation in cells subjected to contact guidance and cyclic strain. Sci Rep 5: 8752. doi: 10.1038/srep08752
|
| [205] | Berg HC (1993) Random Walks in Biology, Princeton: Princeton University Press. |
| [206] |
Köppen M, Fernandez BG, Carvalho L, et al. (2006) Coordinated cell-shape changes control epithelial movement in zebrafish and Drosophila. Development 133: 2671–2681. doi: 10.1242/dev.02439
|
| [207] |
Bretscher MS (2008) On the shape of migrating cells-a 'front-to-back' model. J Cell Sci 121: 2625–2628. doi: 10.1242/jcs.031120
|
| [208] |
Horwitz R, Webb D (2003) Cell migration. Curr Biol 13: R756–R759. doi: 10.1016/j.cub.2003.09.014
|
| [209] | Morris NR (2000) Nuclear migration. From fungi to the mammalian brain. J Cell Biol 148: 1097–1102. |
| [210] |
Petrie R, Doyle AD, Yamada KM (2009) Random versus directionally persistent cell migration. Nat Rev Mol Cell Biol 10: 538–549. doi: 10.1038/nrm2729
|
| [211] |
Cadot B, Gache V, Vasyutina E, et al. (2012) Nuclear movement during myotube formation is microtubule and dynein dependent and is regulated by Cdc42, Par6 and Par3. EMBO Rep 13: 741–749. doi: 10.1038/embor.2012.89
|
| [212] | Lee JSH, Chang MI, Tseng Y, et al. (2005) Cdc42 mediates nucleus movement and MTOC polarization in Swiss 3T3 fibroblasts under mechanical shear stress. Mol Biol Cell 16: 871–880. |
| [213] |
Umeshima H, Hirano T, Kengaku M (2007) Microtubule-based nuclear movement occurs independently of centrosome positioning in migrating neurons. Proc Natl Acad Sci USA 104: 16182–16187. doi: 10.1073/pnas.0708047104
|
| [214] |
Wilson MH, Holzbaur EL (2012) Opposing microtubule motors drive robust nuclear dynamics in developing muscle cells. J Cell Sci 125: 4158–4169. doi: 10.1242/jcs.108688
|
| [215] |
Li R, Gundersen GG (2008) Beyond polymer polarity: how the cytoskeleton builds a polarized cell. Nat Rev Mol Cell Biol 9: 860–873. doi: 10.1038/nrm2522
|
| [216] |
Ostlund C, Folker ES, Choi JC, et al. (2009) Dynamics and molecular interactions of linker of nucleoskeleton and cytoskeleton (LINC) complex proteins. J Cell Sci 122: 4099–4108. doi: 10.1242/jcs.057075
|
| [217] |
Khatau SB, Bloom RJ, Bajpai S, et al. (2012) The distinct roles of the nucleus and nucleus-cytoskeleton connections in three-dimensional cell migration. Sci Rep 2: 488. doi: 10.1038/srep00488
|
| [218] |
Maninova M, Caslavsky J, Vomastek T (2017) The assembly and function of perinuclear actin cap in migrating cells. Protoplasma 254: 1207–1218. doi: 10.1007/s00709-017-1077-0
|
| [219] |
Gay O, Gilquin B, Nakamura F, et al. (2011) RefilinB (FAM101B) targets FilaminA to organize perinuclear actin networks and regulates nuclear shape. Proc Natl Acad Sci USA 108: 11464–11469. doi: 10.1073/pnas.1104211108
|
| [220] |
Capell BC, Collins FS (2006) Human laminopathies: nuclei gone genetically awry. Nat Rev Genet 7: 940–952. doi: 10.1038/nrg1906
|
| [221] |
Khatau SB, Kim DH, Hale CM, et al. (2010) The perinuclear actin cap in health and disease. Nucleus 1: 337–342. doi: 10.4161/nucl.1.4.12331
|
| [222] |
Mogilner A, Keren K (2009) The shape of motile cells. Curr Biol 19: R762–R771. doi: 10.1016/j.cub.2009.06.053
|
| [223] |
Maninova M, Iwanicki MP, Vomastek T (2014) Emerging role for nuclear rotation and orientation in cell migration. Cell Adh Migr 8: 42–48. doi: 10.4161/cam.27761
|
| [224] | Watanabe N, Sekine T, Takagi M, et al. (2005) Deficiency in chromosome congression by the inhibition of Plk1 polo box domain-dependent recognition. J Biol Chem 284: 2344–2353. |
| [225] |
Chung CY, Lee S, Briscoe C, et al. (2000) Role of Rac in controlling the actin cytoskeleton and chemotaxis in motile cells. Proc Natl Acad Sci USA 97: 5225–5230. doi: 10.1073/pnas.97.10.5225
|
| [226] |
Danowski BA, Khodjakov A, Wadsworth P (2001) Centrosome behavior in motile HGF-treated PtK2 cells expressing GFP-gamma tubulin. Cell Motil Cytoskel 50: 59–68. doi: 10.1002/cm.1041
|
| [227] |
Desai RA, Gopal SB, Chen S, et al. (2013) Contact inhibition of locomotion probabilities drive solitary versus collective cell migration. J R Soc Interface 10: 20130717. doi: 10.1098/rsif.2013.0717
|
| [228] |
Niu MY, Mills JC, Nachmias VT (1997) Development of polarity in human erythroleukemia cells: roles of membrane ruffling and the centrosome. Cell Motil Cytoskel 36: 203–215. doi: 10.1002/(SICI)1097-0169(1997)36:3<203::AID-CM1>3.0.CO;2-8
|
| [229] |
Pouthas F, Girard P, Lecaudey V, et al. (2008) In migrating cells, the Golgi complex and the position of the centrosome depend on geometrical constraints of the substratum. J Cell Sci 121: 2406–2414. doi: 10.1242/jcs.026849
|
| [230] |
Ueda M, Gräf R, MacWilliams HK, et al. (1997) Centrosome positioning and directionality of cell movements. Proc Natl Acad Sci USA 94: 9674–9678. doi: 10.1073/pnas.94.18.9674
|
| [231] |
Yvon AM, Walker JW, Danowski B, et al. (2002) Centrosome reorientation in wound-edge cells is cell type specific. Mol Biol Cell 13: 1871–1880. doi: 10.1091/mbc.01-11-0539
|
| [232] |
Jiang X, Bruzewicz DA, Wong AP, et al. (2005) Directing cell migration with asymmetric micropatterns. Proc Natl Acad Sci USA 102: 975–978. doi: 10.1073/pnas.0408954102
|
| [233] |
Hale CM, Chen WC, Khatau SB, et al. (2011) SMRT analysis of MTOC and nuclear positioning reveals the role of EB1 and LIC1 in single-cell polarization. J Cell Sci 124: 4267–4285. doi: 10.1242/jcs.091231
|
| [234] |
Denais CM, Gilbert RM, Isermann P, et al. (2016) Nuclear envelope rupture and repair during cancer cell migration. Science 352: 353–358. doi: 10.1126/science.aad7297
|
| [235] |
Raab M, Gentili M, de Belly H, et al. (2016) ESCRT III repairs nuclear envelope ruptures during cell migration to limit DNA damage and cell death. Science 352: 359–362. doi: 10.1126/science.aad7611
|
| [236] |
Irianto J, Xia Y, Pfeifer CR, et al. (2017) DNA damage follows repair factor depletion and portends genome variation in cancer cells after pore migration. Curr Biol 27: 210–223. doi: 10.1016/j.cub.2016.11.049
|
| [237] |
LeBerre M, Aubertin J, Piel M (2012) Fine control of nuclear confinement identifies a threshold deformation leading to lamina rupture and induction of specific genes. Integr Biol 4: 1406–1414. doi: 10.1039/c2ib20056b
|
| [238] |
Hatch EM, Hetzer MW (2016) Nuclear envelope rupture is induced by actin-based nucleus confinement. J Cell Biol 215: 27–36. doi: 10.1083/jcb.201603053
|
| [239] |
Skau CT, Fischer RS, Gurel P, et al. (2016) FMN2 makes perinuclear actin to protect nuclei during confined migration and promote metastasis. Cell 167: 1571–1585. doi: 10.1016/j.cell.2016.10.023
|
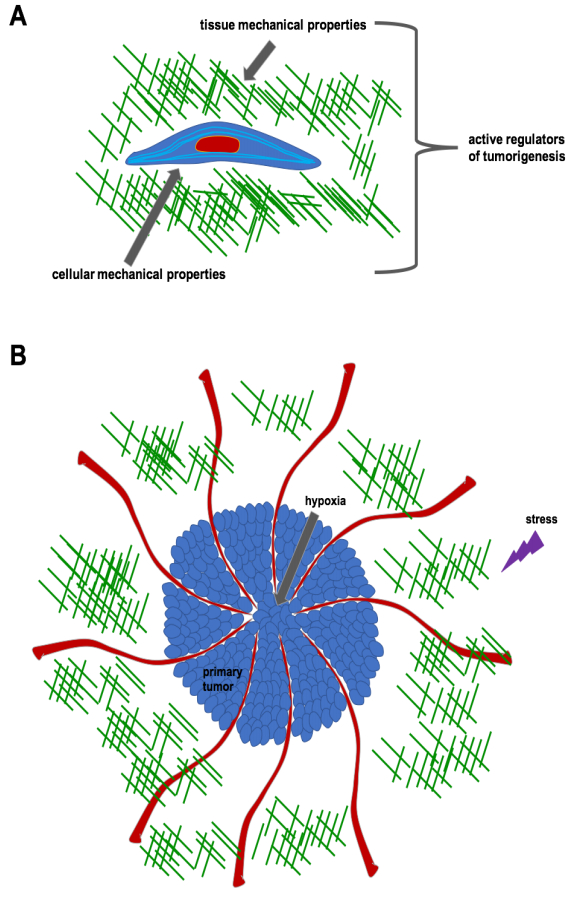
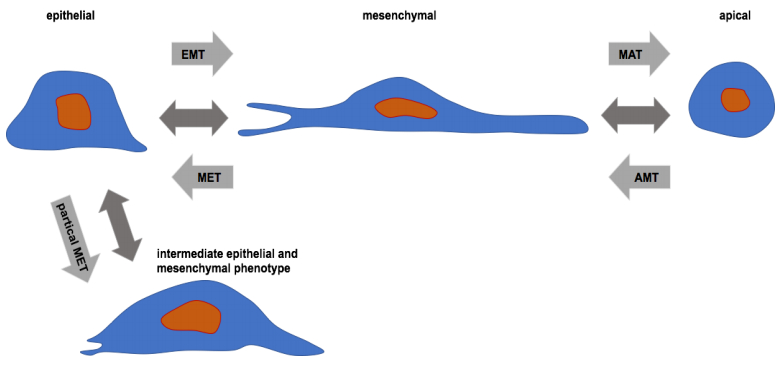
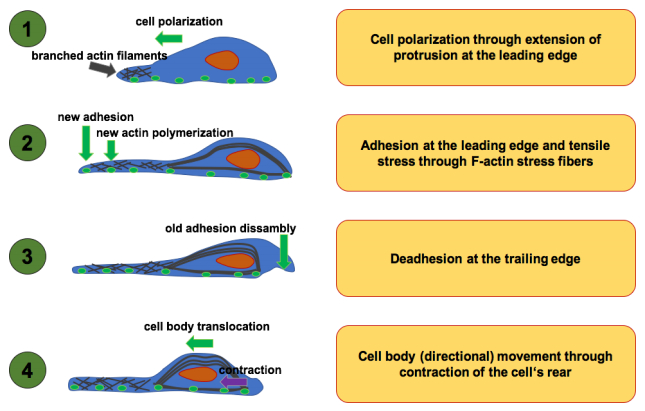
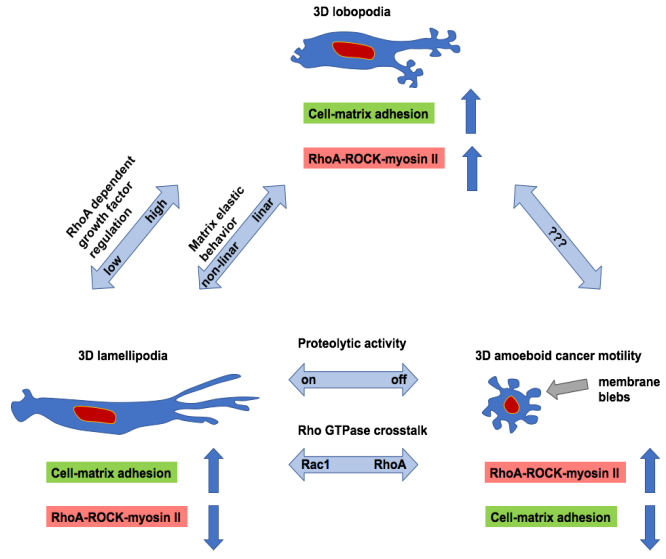
Figures(8)

Claudia Tanja Mierke. Physical role of nuclear and cytoskeletal confinements in cell migration mode selection and switching[J]. AIMS Biophysics, 2017, 4(4): 615-658. doi: 10.3934/biophy.2017.4.615







 DownLoad:
DownLoad: