Citation: Eda Suku, Alejandro Giorgetti. Common evolutionary binding mode of rhodopsin-like GPCRs: Insights from structural bioinformatics[J]. AIMS Biophysics, 2017, 4(4): 543-556. doi: 10.3934/biophy.2017.4.543

| [1] |
Pierce KL, Premont RT, Lefkowitz RJ (2002) Seven-transmembrane receptors. Nat Rev Mol Cell Biol 3: 639–650. doi: 10.1038/nrm908

|
| [2] | Fredriksson R, Lagerström MC, Lundin L, et al. (2003) The G-protein-coupled receptors in the human genome form five main families. Phylogenetic analysis, paralogon groups, and fingerprints. Mol Pharmacol 63: 1256–1272. |
| [3] |
Lagerström MC, Schiöth HB (2008) Structural diversity of G protein-coupled receptors and significance for drug discovery. Nat Rev Drug Discov 7: 339–357. doi: 10.1038/nrd2518
|
| [4] |
Tehan BG, Bortolato A, Blaney, et al. (2014) Unifying family A GPCR theories of activation. Pharmacol Ther 143: 51–60. doi: 10.1016/j.pharmthera.2014.02.004
|
| [5] |
Lans I, Dalton JA, Giraldo J (2015) Helix 3 acts as a conformational hinge in Class A GPCR activation: An analysis of interhelical interaction energies in crystal structures. J Struct Biol 192: 545–553. doi: 10.1016/j.jsb.2015.10.019
|
| [6] |
Trzaskowski B, Latek D, Yuan S, et al. (2012) Action of molecular switches in GPCRs-theoretical and experimental studies. Curr Med Chem 19: 1090–1109. doi: 10.2174/092986712799320556
|
| [7] |
Dalton JA, Lans I, Giraldo J (2015) Quantifying conformational changes in GPCRs: glimpse of a common functional mechanism. BMC Bioinformatics 16: 124. doi: 10.1186/s12859-015-0567-3
|
| [8] |
Katritch V, Fenalti G, Abola EE, et al. (2014) Allosteric sodium in class A GPCR signaling. Trends Biochem Sci 39: 233–244. doi: 10.1016/j.tibs.2014.03.002
|
| [9] |
Krishnan A, Almén MS, Fredriksson R, et al. (2012) The origin of GPCRs: identification of mammalian like Rhodopsin, Adhesion, Glutamate and Frizzled GPCRs in fungi. PLoS One 7: e29817. doi: 10.1371/journal.pone.0029817
|
| [10] |
Willard SS, Koochekpour S (2013) Glutamate, glutamate receptors, and downstream signaling pathways. Int J Biol Sci 9: 948–959. doi: 10.7150/ijbs.6426
|
| [11] |
Springer TA (1990) Adhesion receptors of the immune system. Nature 346: 425–434. doi: 10.1038/346425a0
|
| [12] |
Nichols AS, Floyd DH, Bruinsma SP, et al. (2013) Frizzled receptors signal through G proteins. Cell Signal 25: 1468–1475. doi: 10.1016/j.cellsig.2013.03.009
|
| [13] | Afroze S, Meng F, Jensen K, et al. (2013) The physiological roles of secretin and its receptor. Ann Transl Med 1: 29. |
| [14] |
Costanzi S, Siegel J, Tikhonova IG, et al. (2009) Rhodopsin and the others: a historical perspective on structural studies of G protein-coupled receptors. Curr Pharm Des 15: 3994–4002. doi: 10.2174/138161209789824795
|
| [15] |
Palczewski K, Kumasaka T, Hori T, et al. (2000) Crystal structure of rhodopsin: A G protein-coupled receptor. Science 289: 739–745. doi: 10.1126/science.289.5480.739
|
| [16] |
Isberg V, Mordalski S, Munk C, et al. (2016) GPCRdb: an information system for G protein-coupled receptors. Nucleic Acids Res 44: D356–D364. doi: 10.1093/nar/gkv1178
|
| [17] |
Zhang H, Qiao A, Yang D, et al. (2017) Structure of the full-length glucagon class B G-protein-coupled receptor. Nature 546: 259–264. doi: 10.1038/nature22363
|
| [18] |
Lundstrom K (2006) Latest development in drug discovery on G protein-coupled receptors. Curr Protein Pept Sci 7: 465–470. doi: 10.2174/138920306778559403
|
| [19] |
Wacker D, Stevens RC, Roth BL (2017) How ligands illuminate GPCR molecular pharmacology. Cell 170: 414–427. doi: 10.1016/j.cell.2017.07.009
|
| [20] |
Moreira IS (2014) Structural features of the G-protein/GPCR interactions. Biochim Biophys Acta 1840: 16–33. doi: 10.1016/j.bbagen.2013.08.027
|
| [21] |
Venkatakrishnan AJ, Deupi X, Lebon G, et al. (2013) Molecular signatures of G-protein-coupled receptors. Nature 494: 185–194. doi: 10.1038/nature11896
|
| [22] |
Katritch V, Cherezov V, Stevens RC (2013) Structure-function of the G protein-coupled receptor superfamily. Annu Rev Pharmacol Toxicol 53: 531–556. doi: 10.1146/annurev-pharmtox-032112-135923
|
| [23] |
Wolf S, Grünewald S (2015) Sequence, structure and ligand binding evolution of rhodopsin-like G protein-coupled receptors: a crystal structure-based phylogenetic analysis. PLoS One 10: e0123533. doi: 10.1371/journal.pone.0123533
|
| [24] |
Kratochwil NA, Malherbe P, Lindemann L, et al. (2005) An automated system for the analysis of G protein-coupled receptor transmembrane binding pockets: alignment, receptor-based pharmacophores, and their application. J Chem Inf Model 45: 1324–1336. doi: 10.1021/ci050221u
|
| [25] |
Rovati GE, Capra V, Neubig RR (2007) The highly conserved DRY motif of class A G protein-coupled receptors: beyond the ground state. Mol Pharmacol 71: 959–964. doi: 10.1124/mol.106.029470
|
| [26] |
Urizar E, Claeysen S, Deupí X, et al. (2005) An activation switch in the rhodopsin family of G protein-coupled receptors: the thyrotropin receptor. J Biol Chem 280: 17135–17141. doi: 10.1074/jbc.M414678200
|
| [27] |
Caltabiano G, Gonzalez A, Cordomí A, et al. (2013) The role of hydrophobic amino acids in the structure and function of the rhodopsin family of G protein-coupled receptors. Methods Enzymol 520: 99–115. doi: 10.1016/B978-0-12-391861-1.00005-8
|
| [28] |
Wheatley M, Wootten D, Conner MT, et al. (2012) Lifting the lid on GPCRs: the role of extracellular loops. Br J Pharmacol 165: 1688–1703. doi: 10.1111/j.1476-5381.2011.01629.x
|
| [29] |
Zhang D, Zhao Q, Wu B (2015) Structural studies of G protein-coupled receptors. Mol Cells 38: 836–842. doi: 10.14348/molcells.2015.0263
|
| [30] | Ziarek JJ, Kleist AB, London N, et al. (2017) Structural basis for chemokine recognition by a G protein-coupled receptor and implications for receptor activation. Sci Signal 10: 471. |
| [31] |
Stansfeld PJ, Goose JE, Caffrey M, et al. (2015) MemProtMD: automated insertion of membrane protein structures into explicit lipid membranes. Structure 23: 1350–1361. doi: 10.1016/j.str.2015.05.006
|
| [32] |
Rose PW, Prlić A, Altunkaya A, et al. (2017) The RCSB protein data bank: integrative view of protein, gene and 3D structural information. Nucleic Acids Res 45: D271–D281. doi: 10.1093/nar/gkw1042
|
| [33] | Vangone A, Bonvin AM (2015) Contacts-based prediction of binding affinity in protein-protein complexes. Elife 4: e07454. |
| [34] |
Venkatakrishnan AJ, Deupi X, Lebon G, et al. (2016) Diverse activation pathways in class A GPCRs converge near the G-protein-coupling region. Nature 536: 484–487. doi: 10.1038/nature19107
|
| [35] |
Crooks GE, Hon G, Chandonia JM, et al. (2004) WebLogo: a sequence logo generator. Genome Res 14: 1188–1190. doi: 10.1101/gr.849004
|
| [36] |
Simonetti FL, Teppa E, Chernomoretz A, et al. (2013) MISTIC: Mutual information server to infer coevolution. Nucleic Acids Res 41: W8–W14. doi: 10.1093/nar/gkt427
|
| [37] |
Gloor GB, Martin LC, Wahl LM, et al. (2005) Mutual information in protein multiple sequence alignments reveals two classes of coevolving positions. Biochemistry 44: 7156–7165. doi: 10.1021/bi050293e
|
| [38] |
Martin LC, Gloor GB, Dunn SD, et al. (2005) Using information theory to search for co-evolving residues in proteins. Bioinformatics 21: 4116–4124. doi: 10.1093/bioinformatics/bti671
|
| [39] | Ward Jr, Joe H (1963) Hierarchical grouping to optimize an objective function. J Am Stat Assoc 301: 236–244. |
| [40] |
Irwin JJ, Shoichet BK (2005) ZINC-a free database of commercially available compounds for virtual screening. J Chem Inf Model 45: 177–182. doi: 10.1021/ci049714+
|
| [41] |
Chan WK, Zhang H, Yang J, et al. (2015) GLASS: a comprehensive database for experimentally validated GPCR-ligand associations. Bioinformatics 31: 3035–3042. doi: 10.1093/bioinformatics/btv302
|
| [42] |
Lee SM, Booe JM, Pioszak AA (2015) Structural insights into ligand recognition and selectivity for classes A, B, and C GPCRs. Eur J Pharmacol 763: 196–205. doi: 10.1016/j.ejphar.2015.05.013
|
| [43] |
Buslje CM, Santos J, Delfino JM, et al. (2009) Correction for phylogeny, small number of observations and data redundancy improves the identification of coevolving amino acid pairs using mutual information. Bioinformatics 25: 1125–1131. doi: 10.1093/bioinformatics/btp135
|
| [44] |
Marco E, Foucaud M, Langer I, et al. (2007) Mechanism of activation of a G protein-coupled receptor, the human cholecystokinin-2 receptor. J Biol Chem 282: 28779–28790. doi: 10.1074/jbc.M700349200
|
| [45] |
Kooistra AJ, Kuhne S, Esch IJP, et al. (2013) A structural chemogenomics analysis of aminergic GPCRs: lessons for histamine receptor ligand design. Br J Pharmacol 170: 101–126. doi: 10.1111/bph.12248
|
| [46] |
Regard JB, Sato IT, Coughlin SR (2008) Anatomical profiling of G protein-coupled receptor expression. Cell 135: 561–571. doi: 10.1016/j.cell.2008.08.040
|
| [47] | Stevens RC, Cherezov V, Katritch V, et al. (2013) The GPCR Network: a large-scale collaboration to determine human GPCR structure and function. Nat Rev Drug Discov 12: 25–34. |
| [48] | Sanger GJ, Furness JB (2016) Ghrelin and motilin receptors as drug targets for gastrointestinal disorders. Nat Rev Gastroenterol Hepatol 13: 38–48. |
| [49] |
Elphick MR (1998) An invertebrate G-protein coupled receptor is a chimeric cannabinoid/melanocortin receptor. Brain Res 780: 170–173. doi: 10.1016/S0006-8993(97)01297-3
|
| [50] |
Harikumar KG, Puri V, Singh RD, et al. (2005) Differential effects of modification of membrane cholesterol and sphingolipids on the conformation, function, and trafficking of the G protein-coupled cholecystokinin receptor. J Biol Chem 280: 2176–2185. doi: 10.1074/jbc.M410385200
|
| [51] |
Bhalla S, Zhang Z, Patterson N, et al. (2010) Effect of endothelin-A receptor antagonist on mu, delta and kappa opioid receptor-mediated antinociception in mice. Eur J Pharmacol 635: 62–71. doi: 10.1016/j.ejphar.2010.03.003
|
| [52] |
Gao L, Yu LC (2004) Involvement of opioid receptors in the oxytocin-induced antinociception in the central nervous system of rats. Regul Pept 120: 53–58. doi: 10.1016/j.regpep.2004.02.011
|
| [53] |
Deupi X, Standfuss J (2011) Structural insights into agonist-induced activation of G-protein-coupled receptors. Curr Opin Struct Biol 21: 541–551. doi: 10.1016/j.sbi.2011.06.002
|
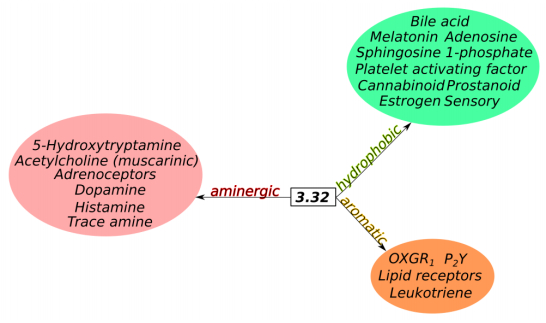
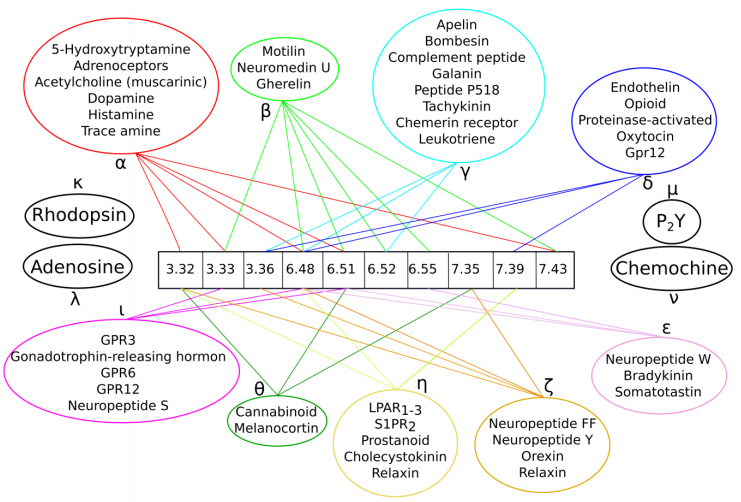
Figures(6) / Tables(1)

Eda Suku, Alejandro Giorgetti. Common evolutionary binding mode of rhodopsin-like GPCRs: Insights from structural bioinformatics[J]. AIMS Biophysics, 2017, 4(4): 543-556. doi: 10.3934/biophy.2017.4.543







 DownLoad:
DownLoad: