Citation: Ekaterina Bereznyak, Natalia Gladkovskaya, Evgeniy Dukhopelnykov, Anastasiya Gerus, Anastasiya Lantushenko, Maxim Evstigneev. Thermal analysis of ligand-DNA interaction: determination of binding parameters[J]. AIMS Biophysics, 2015, 2(4): 423-440. doi: 10.3934/biophy.2015.4.423

| [1] |
Finley A, Copeland RA (2014) Small molecule control of chromatin remodeling. Chem Biol 21: 1196–1210. doi: 10.1016/j.chembiol.2014.07.024

|
| [2] |
Prinjha R, Tarakhovsky A (2013) Chromatin targeting drugs in cancer and immunity. Genes Dev 27: 1731–1738. doi: 10.1101/gad.221895.113
|
| [3] |
Rabbani A, Finn RM, Ausio J (2005) The anthracycline antibiotics: antitumor drugs that alter chromatin structure. Bioessays 27: 50–56. doi: 10.1002/bies.20160
|
| [4] |
Majumder P, Pradhan SK, Devi PG, et al. (2007) Chromatin as a target for the DNA-binding anticancer drugs. Subcell Biochem 41: 145–189. doi: 10.1007/1-4020-5466-1_8
|
| [5] |
Banerjee A, Majumder P, Sanyal S, et al. (2014) The DNA intercalators ethidium bromide and propidium iodide also bind to core histones. FEBS Open Bio 4: 251–259. doi: 10.1016/j.fob.2014.02.006
|
| [6] |
Hajihassan Z, Rabbani-Chadegani A (2009) Studies on the binding affinity of anticancer drug mitoxantrone to chromatin, DNA and histone proteins. J Biomed Sci 16: 31. doi: 10.1186/1423-0127-16-31
|
| [7] |
Rabbani-Chadegani A, Chamani E, Hajihassan Z (2009) The effect of vinca alkaloid anticancer drug, vinorelbine, on chromatin and histone proteins in solution. Eur J Pharmacol 613: 34–38. doi: 10.1016/j.ejphar.2009.04.040
|
| [8] |
Rabbani-Chadegani A, Keyvani-Ghamsari S, Zarkar N (2011) Spectroscopic studies of dactinomycin and vinorelbine binding to deoxyribonucleic acid and chromatin. Spectrochim Acta A Mol Biomol Spectrosc 84: 62–67. doi: 10.1016/j.saa.2011.08.064
|
| [9] |
Rabbani A, Iskandar M, Ausio J (1999) Daunomycin-induced unfolding and aggregation of chromatin. J Biol Chem 274: 18401–18406. doi: 10.1074/jbc.274.26.18401
|
| [10] |
Mir MA, Majee S, Das S, et al. (2003) Association of chromatin with anticancer antibiotics, mithramycin and chromomycin A3. Bioorg Med Chem 11: 2791–2801. doi: 10.1016/S0968-0896(03)00211-6
|
| [11] |
Mir MA, Dasgupta D (2001) Association of the anticancer antibiotic chromomycin A(3) with the nucleosome: role of core histone tail domains in the binding process. Biochemistry 40: 11578–11585. doi: 10.1021/bi010731r
|
| [12] |
Mir MA, Dasgupta D (2001) Interaction of antitumor drug, mithramycin, with chromatin. Biochem Biophys Res Commun 280: 68–74. doi: 10.1006/bbrc.2000.4075
|
| [13] |
Hagmar P, Pierrou S, Nielsen P, et al. (1992) Ionic strength dependence of the binding of methylene blue to chromatin and calf thymus DNA. J Biomol Struct Dyn 9: 667–679. doi: 10.1080/07391102.1992.10507947
|
| [14] |
Mir MA, Das S, Dasgupta D (2004) N-terminal tail domains of core histones in nucleosome block the access of anticancer drugs, mithramycin and daunomycin, to the nucleosomal DNA. Biophys Chem 109: 121–135. doi: 10.1016/j.bpc.2003.10.023
|
| [15] |
Hurley LH (2002) DNA and its associated processes as targets forcancer therapy. Nat Rev Cancer 2: 188–200. doi: 10.1038/nrc749
|
| [16] | Veselkov AN, Davies DB (2002) Anticancer drug design, Sevastopol: Sevntu press, 259. |
| [17] | Veselkov AN, Maleev VYa, Glibin EN, et al. (2003) Structure–activity relation for synthetic phenoxazone drugs.Evidence for a direct correlation between DNA binding and pro-apoptotic activity. Eur J Biochem 270: 4200–4207 |
| [18] |
Andersson J, Lincoln P (2011) Stereoselectivity for DNA threading intercalation of short binuclear ruthenium complexes. J Phys Chem B 115: 14768–14775. doi: 10.1021/jp2062767
|
| [19] | Chaurasiya KR, Paramanathan T, McCauley MJ, et al. (2010) Biophysical characterization of DNA binding from single molecule force measurements. Phys Life Rev 7: 299–341. |
| [20] |
Nordell P, Lincoln P (2005) Mechanism of DNA threading intercalation of binuclear Ru complexes: uni- or bimolecular pathways depending on ligand structure and binding density. J Am Chem Soc 127: 9670–9671. doi: 10.1021/ja0521674
|
| [21] | Andersson J, Li M, Lincoln P (2010) AT-specific DNA binding of binuclear ruthenium complexes at the border of threading intercalation. Chemistry 16: 11037–11046. |
| [22] | Wilhelmsson LM, Lincoln P, Nordґen B (2006) Slow DNA binding. In: Waring M, editor. Sequence-Specific DNA Binding Agents, The Royal Society of Chemistry, Cambridge, 69–95. |
| [23] |
Palchaudhuri R, Hergenrother PJ (2007) DNA as a target for anticancer compounds: methods to determine the mode of binding and the mechanism of action. Curr Opin Biotechnol 18: 497–503. doi: 10.1016/j.copbio.2007.09.006
|
| [24] | Barton TF, Cooney RP, Denny WA (1992) Surface-enhanced Raman spectroscopic study of amsacrine and amsacrine–DNA interactions. J Raman Spectrosc 23: 341–345. |
| [25] |
Rodger A, Blagbrough IS, Adlam G, et al. (1994) DNA binding of a spermine derivative: Spectroscopic study of anthracene-9-carbonyl-n1-spermine with poly[d(G-C)·(d(G-C))] and poly[d(A-T) · d(A-T)]. Biopolymers 34: 1583–1593. doi: 10.1002/bip.360341203
|
| [26] | Hackl EV, Galkin VL, Blagoi YP (2004) DNA interaction with biologically active divalent metal ions: binding constants calculation. Int J Biol Macromol 34: 303–308. |
| [27] |
Evstigneev MP, Mykhina YV, Davies DB (2005) Complexation of daunomycin with a DNA oligomer in the presence of an aromatic vitamin (B2) determined by NMR spectroscopy. Biophys Chem 118: 118–127. doi: 10.1016/j.bpc.2005.08.007
|
| [28] |
Hackl EV, Blagoi YP (2005) The effect of temperature on DNA structural transitions under the action of Cu2+ and Ca2+ ions in aqueous solutions. Biopolymers 77: 315–324. doi: 10.1002/bip.20225
|
| [29] | Kruglova EB, Gladkovskaia NA, Maleev V (2005) The use of the spectrophotometric analysis for the calculation of the thermodynamic parameters in actinocin derivative-DNA systems. Biofizika 50: 253–264. |
| [30] | Evstigneev MP, Baranovskii SF, Rybakova KA, et al. (2006) 1H NMR study of the complexation of the quinolone antibiotic norfloxacin with DNA. Mol Biol (Mosk) 40: 894–899. |
| [31] |
Evstigneev MP, Rybakova KA, Davies DB (2006) Complexation of norfloxacin with DNA in the presence of caffeine. Biophys Chem 121: 84–95. doi: 10.1016/j.bpc.2005.12.003
|
| [32] |
Williams AK, Dasilva SC, Bhatta A, et al. (2012) Determination of the drug-DNA binding modes using fluorescence-based assays. Anal Biochem 422: 66–73. doi: 10.1016/j.ab.2011.12.041
|
| [33] |
Anupama B, Sunita M, Shiva LD, et al. (2014) Synthesis, spectral characterization, DNA binding studies and antimicrobial activity of Co(II), Ni(II), Zn(II), Fe(III) and VO(IV) complexes with 4-aminoantipyrine Schiff base of ortho-vanillin. J Fluoresc 24: 1067–1076. doi: 10.1007/s10895-014-1386-z
|
| [34] | Zasedatelev AS, Gurskii GV, Vol'kenshtein MV (1971) Theory of one-dimensional adsorption. I. Adsorption of small molecules on a homopolymer. Molecular biology 5: 194–198. |
| [35] |
McGhee JD, von Hippel PH (1974) Theoretical aspects of DNA-protein interactions: co-operative and non-co-operative binding of large ligands to a one-dimensional homogeneous lattice. J Mol Biol 86: 469–489. doi: 10.1016/0022-2836(74)90031-X
|
| [36] |
Schellman JA (1974) Cooperative Multisite Binding to DNA. Israel Journal of Chemistry 12: 219–238. doi: 10.1002/ijch.197400021
|
| [37] |
Nechipurenko YD, Gursky GV (1986) Cooperative effects on binding of proteins to DNA. Biophys Chem 24: 195–209. doi: 10.1016/0301-4622(86)85025-6
|
| [38] |
Lando DY, Teif VB (2000) Long-range interactions between ligands bound to a DNA molecule give rise to adsorption with the character of phase transition of the first kind. J Biomol Struct Dyn 17: 903–911. doi: 10.1080/07391102.2000.10506578
|
| [39] |
Teif VB, Rippe K (2010) Statistical-mechanical lattice models for protein-DNA binding in chromatin. J Phys Condens Matter 22: 414105. doi: 10.1088/0953-8984/22/41/414105
|
| [40] | Breslauer KJ, Freire E, Straume M (1992) Calorimetry: a tool for DNA and ligand-DNA studies. Methods Enzymol 211: 533–567. |
| [41] |
Doyle ML (1997) Characterization of binding interactions by isothermal titration calorimetry. Curr Opin Biotechnol 8: 31–35. doi: 10.1016/S0958-1669(97)80154-1
|
| [42] | Jelesarov I, Bosshard HR (1999) Isothermal titration calorimetry and differential scanning calorimetry as complementary tools to investigate the energetics of biomolecular recognition. J Mol Recognit 12: 3–18. |
| [43] | Haq I, Chowdhry BZ, Jenkins TC (2001)Calorimetric techniques in the study of high-order DNA-drug interactions. Methods Enzymol 340: 109–149. |
| [44] | Thomson JA, Ladbury JE (2004) Isothermal titration calorimetry: a tutorial. In Ladbury JE, Doyle ML, editors. Biocalorimetry 2. Applications of Calorimetry in the Biological Sciences, Chichester: John Wiley & Sons, 35–58. |
| [45] | Holdgate GA, Ward WH (2005) Measurements of binding thermodynamics in drug discovery. Drug Discov Today 10: 1543–1550. |
| [46] |
Freyer MW, Lewis EA (2008) Isothermal titration calorimetry: experimental design, data analysis, and probing macromolecule/ligand binding and kinetic interactions. Methods Cell Biol 84: 79–113. doi: 10.1016/S0091-679X(07)84004-0
|
| [47] |
Bhadra K, Maiti M, Kumar GS (2008) Berberine–DNA complexation: New insights into the cooperative binding and energetic aspects. Biochimica et Biophysica Acta 1780: 1054–1061. doi: 10.1016/j.bbagen.2008.05.005
|
| [48] | Hossain M, Kumar GS (2009) DNA intercalation of methylene blue and quinacrine: new insights into base and sequence specificity from structural and thermodynamic studies with polynucleotides. Mol Biosyst 5: 1311–1322. |
| [49] |
Crane-Robinson C, Dragan AI, Read CM (2009) Defining the thermodynamics of protein/DNA complexes and their components using micro-calorimetry. Methods Mol Biol 543: 625–651. doi: 10.1007/978-1-60327-015-1_37
|
| [50] |
Kabir A, Kumar GS (2013) Binding of the biogenic polyamines to deoxyribonucleic acids of varying base composition: base specificity and the associated energetics of the interaction. PLoS One 8: e70510. doi: 10.1371/journal.pone.0070510
|
| [51] |
Kumar S, Spano MN, Arya DP (2014) Shape readout of AT-rich DNA by carbohydrates. Biopolymers 101: 720–732. doi: 10.1002/bip.22448
|
| [52] |
Basu A, Kumar GS (2015) Thermodynamic characterization of proflavine–DNA binding through microcalorimetric studies. J Chem Thermodyn 87: 1–7. doi: 10.1016/j.jct.2015.03.009
|
| [53] | Basu A, Kumar GS (2015) Studies on the interaction of the food colorant tartrazine with double stranded deoxyribonucleic acid. J Biomol Struct Dyn 10: 1–8. |
| [54] | Chaires JB (1997) Possible origin of differences between van't Hoff and calorimetric enthalpy estimates. Biophys Chem 64: 15–23. |
| [55] | Chaires JB (2008) Calorimetry and thermodynamics in drug design. Annu Rev Biophys 37: 135–151. |
| [56] |
Janjua NK, Siddiqa A, Yaqub A, et al. (2009) Spectrophotometric analysis of flavonoid-DNA binding interactions at physiological conditions. Spectrochim Acta A Mol Biomol Spectrosc 74: 1135–1137. doi: 10.1016/j.saa.2009.09.022
|
| [57] |
Temerk YM, Ibrahim MS, Kotb M (2009) Voltammetric and spectroscopic studies on binding of antitumor Morin, Morin-Cu complex and Morin-beta-cyclodextrin with DNA. Spectrochim Acta A Mol Biomol Spectrosc 71: 1830–1836. doi: 10.1016/j.saa.2008.07.001
|
| [58] |
Baranovskii SF, Chernyshev DN, Buchel’nikov AS, et al. (2011) Thermodynamic analysis of complex formation of ethidium bromide with DNA in water solutions. Biophysics 56: 214–219. doi: 10.1134/S0006350911020023
|
| [59] | Chaires JB (1997) Energetics of drug-DNA interactions. Biopolymers 44: 201–215. |
| [60] |
Ren J, Jenkins TC, Chaires JB (2000) Energetics of DNA intercalation reactions. Biochemistry 39: 8439–8447. doi: 10.1021/bi000474a
|
| [61] |
Davies DB, Veselkov AN (1996) Structural and thermodynamical analysis of molecular complexation by 1H NMR spectroscopy. Intercalation of ethidium bromide with the isomeric deoxytetranucleoside triphosphates 5’-d(GpCpGpC) and 5’-d(CpGpCpG) in aqueous solution. J Chem Soc Faraday Trans 92: 3545-3557. doi: 10.1039/ft9969203545
|
| [62] |
Kostjukov VV, Pahomov VI, Andrejuk DD, et al. (2007) Investigation of the complexation of the anti-cancer drug novantrone with the hairpin structure of the deoxyheptanucleotide 5′-d(GpCpGpApApGpC). J Mol Struct 843: 78–86. doi: 10.1016/j.molstruc.2006.12.036
|
| [63] |
Wartell RM, Benight AS (1985) Thermal denaturation of DNA molecules: A comparison of theory with experiment. Phys Rep 126: 67–107. doi: 10.1016/0370-1573(85)90060-2
|
| [64] |
Rice SA, Doty P (1957) The Thermal Denaturation of Desoxyribose Nucleic Acid. J Am Chem Soc 79: 3937–3947. doi: 10.1021/ja01572a001
|
| [65] |
Guedin A, Lacroix L, Mergny JL (2010) Thermal melting studies of ligand DNA interactions. Methods Mol Biol 613: 25–35. doi: 10.1007/978-1-60327-418-0_2
|
| [66] | Goldstein G, Stern KG (1950) Experiments on the sonic, thermal, and enzymic depolymerization of desoxyribosenucleic acid. J Struc Chem 5: 687–708. |
| [67] |
Thomas R (1954) Recherches sur la d'enaturation des acides desoxyribonucléiques. Biochimica et Biophysica Acta 14: 231–240. doi: 10.1016/0006-3002(54)90163-8
|
| [68] | Frank-Kamenetskii M (1965) Theory of the helix–coil transition for deoxyribonucleic acids with additional connections between the chains. Vysokomolekulyarnye Soedineniya 7: 354–361. |
| [69] | Frank-Kamenetskii M (1968) Consideration of helix-coil transition in homopolymers by the most probable distribution method. Mol Biol 2: 408–419. |
| [70] |
Stewart CR (1968) Broadening by acridine orange of the thermal transition of DNA. Biopolymers 6: 1737–1743. doi: 10.1002/bip.1968.360061208
|
| [71] |
Lazurkin YS, Frank-Kamenetskii MD, Trifonov EN (1970) Melting of DNA: its study and application as a research method. Biopolymers 9: 1253–1306. doi: 10.1002/bip.1970.360091102
|
| [72] |
Barcelo F, Capo D, Portugal J (2002) Thermodynamic characterization of the multivalent binding of chartreusin to DNA. Nucleic Acids Res 30: 4567–4573. doi: 10.1093/nar/gkf558
|
| [73] |
Zhong W, Yu JS, Liang Y (2003) Chlorobenzylidine-herring sperm DNA interaction: binding mode and thermodynamic studies. Spectrochim Acta A Mol Biomol Spectrosc 59: 1281–1288. doi: 10.1016/S1386-1425(02)00301-3
|
| [74] |
Vardevanyan PO, Antonyan AP, Hambardzumyan LA, et al. (2013) Thermodynamic analysis of DNA complexes with methylene blue, ethidium bromide and Hoechst 33258. Biopolym. Cell 29: 515–520. doi: 10.7124/bc.000843
|
| [75] | Hajian R, Guan Huat T (2013) Spectrophotometric Studies on the Thermodynamics of the ds-DNA Interaction with Irinotecan for a Better Understanding of Anticancer Drug-DNA Interactions. J Spectrosc 2013: 1–8. |
| [76] | Cooper A, Johnson CM (1994) Introduction to microcalorimetry and biomolecular energetics. Methods Mol Biol 22: 109–24. |
| [77] | Rosgen J, Hinz HJ (1999) Theory and practice of DSC mesuarements on proteins. In: Kemp RB, editor. Handbook of Thermal Analysis and Calorimetry, Vol.4, From Macromolecules to Man, Amsterdam: Elsevier, 63–108. |
| [78] |
Bruylants G, Wouters J, Michaux C (2005) Differential scanning calorimetry in life science: thermodynamics, stability, molecular recognition and application in drug design. Curr Med Chem 12: 2011–2020. doi: 10.2174/0929867054546564
|
| [79] |
Spink CH (2008) Differential scanning calorimetry. Methods Cell Biol 84: 115–141. doi: 10.1016/S0091-679X(07)84005-2
|
| [80] |
Bereznyak EG, Gladkovskaya NA, Khrebtova AS, et al. (2009) Peculiarities of DNA-proflavine binding under different concentration ratios. Biophysics 54: 574–580. doi: 10.1134/S0006350909050030
|
| [81] | Garbett N (2011) The Use of Calorimetry to Study Ligand–DNA Interactions. In: Aldrich-Wright J, editor. Metallointercalators, Vienna: Springer, 299–324. |
| [82] |
Ising E (1925) Beitrag zur Theorie des Ferromagnetismus. Zeitschrift für Physik 31: 253–258. doi: 10.1007/BF02980577
|
| [83] | Poland DC, Scheraga HA (1970) The theory of helix coil transition, New York: Academic Press. |
| [84] |
Peyrard M, Bishop AR (1989) Statistical mechanics of a nonlinear model for DNA denaturation. Phys Rev Lett 62: 2755–2758. doi: 10.1103/PhysRevLett.62.2755
|
| [85] |
Dauxois T, Peyrard M, Bishop AR (1993) Entropy-driven DNA denaturation. Physical Review E 47: R44–R47. doi: 10.1103/PhysRevE.47.R44
|
| [86] | Dauxois T, Peyrard M, Bishop AR (1993) Dynamics and thermodynamics of a nonlinear model for DNA denaturation. Phys Rev E Stat Phys Plasmas Fluids Relat Interdiscip Topics 47: 684–695. |
| [87] | Grosberg AIU, Khokhlov AR (1994) Statistical physics of macromolecules, New York: AIP Press. |
| [88] |
Frank-Kamenetskii MD, Prakash S (2014) Fluctuations in the DNA double helix: A critical review. Phys Life Rev 11: 153–170. doi: 10.1016/j.plrev.2014.01.005
|
| [89] | Cantor CR, Schimmel PR (1980) Biophysical chemistry. 3. The behavior of biological macromolecules, San Francisco, Freeman. |
| [90] |
Zimm BH, Bragg JK (1959) Theory of the Phase Transition between Helix and Random Coil in Polypeptide Chains. J Chem Phys 31: 526–535. doi: 10.1063/1.1730390
|
| [91] |
Crothers DM (1971) Statistical thermodynamics of nucleic acid melting transitions with coupled binding equilibria. Biopolymers 10: 2147–2160. doi: 10.1002/bip.360101110
|
| [92] |
McGhee JD (1976) Theoretical calculations of the helix-coil transition of DNA in the presence of large, cooperatively binding ligands. Biopolymers 15: 1345–1375. doi: 10.1002/bip.1976.360150710
|
| [93] |
Akhrem AA, Fridman AS, Lando D (1985) Theory of helix-coil transition of the heterogeneous DNA-heteroqeneous ligands complexes. Biopolym Cell 1: 171–179. doi: 10.7124/bc.00017E
|
| [94] |
Lando D (1994) A theoretical consideration of the influence of selective binding of small ligands on DNA helix-coil transition. J Biomol Struct Dyn 12: 343–354. doi: 10.1080/07391102.1994.10508744
|
| [95] | Akhrem AA, Lando D (1979) Influence of ligands characteristic of selective binding to a certain type of base pairs on DNA helix-coil transition I. Model. Theory. Mol Biol (Mosk) 13: 1098–1109. |
| [96] | Akhrem AA, Lando D, Shpakovskii AG, et al. (1990) The effect of long-range interactions between adsorbed ligands on the DNA helix-coil transition. Mol Biol (Mosk) 24: 649–656. |
| [97] | Lando D, Ivanova MA, Akhrem AA (1980) Effect of changes in the stoichiometry of DNA-ligand complexes during heat denaturation of DNA on helix-coil transition parameters. Mol Biol (Mosk) 14: 1281–1288. |
| [98] |
Karapetian AT, Mehrabian NM, Terzikian GA, et al. (1996) Theoretical treatment of melting of complexes of DNA with ligands having several types of binding sites on helical and single-stranded DNA. J Biomol Struct Dyn 14: 275–283. doi: 10.1080/07391102.1996.10508118
|
| [99] |
Plum GE, Bloomfield VA (1990) Structural and electrostatic effects on binding of trivalent cations to double-stranded and single-stranded poly[d (AT)]. Biopolymers 29: 13–27. doi: 10.1002/bip.360290105
|
| [100] |
Spink CH, Chaires JB (1997) Thermodynamics of the Binding of a Cationic Lipid to DNA. J Am Chem Soc 119: 10920–10928. doi: 10.1021/ja964324s
|
| [101] |
Leng F, Chaires JB, Waring MJ (2003) Energetics of echinomycin binding to DNA. Nucleic Acids Res 31: 6191–6197. doi: 10.1093/nar/gkg826
|
| [102] |
Pasic L, Sepcic K, Turk T, et al. (2001) Characterization of parazoanthoxanthin A binding to a series of natural and synthetic host DNA duplexes. Arch Biochem Biophys 393: 132–142. doi: 10.1006/abbi.2001.2469
|
| [103] |
Portugal J, Cashman DJ, Trent JO, et al. (2005) A new bisintercalating anthracycline with picomolar DNA binding affinity. J Med Chem 48: 8209–8219. doi: 10.1021/jm050902g
|
| [104] |
Liu Y-J, Wei X, Mei W-J, et al. (2007) Synthesis, characterization and DNA binding studies of ruthenium(II) complexes: [Ru(bpy)2(dtmi)]2+ and [Ru(bpy)2(dtni)]2+. Transit Metal Chem 32: 762–768. doi: 10.1007/s11243-007-0246-y
|
| [105] |
Peng B, Chen X, Du KJ, et al. (2009) Synthesis, characterization and DNA-binding studies of ruthenium(II) mixed-ligand complexes containing dipyrido[1,2,5]oxadiazolo[3,4-b]quinoxaline. Spectrochim Acta A Mol Biomol Spectrosc 74: 896–901. doi: 10.1016/j.saa.2009.08.031
|
| [106] | Barcelo F, Portugal J (2004) Elsamicin A binding to DNA. A comparative thermodynamic characterization. FEBS Lett 576: 68–72. |
| [107] |
Barcelo F, Scotta C, Ortiz-Lombardia M, et al. (2007) Entropically-driven binding of mithramycin in the minor groove of C/G-rich DNA sequences. Nucleic Acids Res 35: 2215–2226. doi: 10.1093/nar/gkm037
|
| [108] | Marky LA, Blumenfeld KS, Breslauer KJ (1983) Calorimetric and spectroscopic investigation of drug-DNA interactions. I. The binding of netropsin to poly d(AT). Nucleic Acids Res 11: 2857–2870. |
| [109] |
Remeta DP, Mudd CP, Berger RL, et al (1993) Thermodynamic characterization of daunomycin-DNA interactions: comparison of complete binding profiles for a series of DNA host duplexes. Biochemistry 32: 5064–5073. doi: 10.1021/bi00070a014
|
| [110] | Brandts JF, Lin LN (1990) Study of strong to ultratight protein interactions using differential scanning calorimetry. Biochemistry 29: 6927–6940. |
| [111] | Barone G, Catanzano F, Del Vecchio P, et al. (1995) Differential scanning calorimetry as a tool to study protein-ligand interactions. Pure Appl Chem 67: 1867–1872. |
| [112] |
Dassie SA, Celej MS, Fidelio GD (2005) Protein Unfolding Coupled to Ligand Binding: Differential Scanning Calorimetry Simulation Approach. J Chem Educ 82: 85. doi: 10.1021/ed082p85
|
| [113] |
Celej MS, Dassie SA, Gonzalez M, et al. (2006) Differential scanning calorimetry as a tool to estimate binding parameters in multiligand binding proteins. Anal Biochem 350: 277–284. doi: 10.1016/j.ab.2005.12.029
|
| [114] |
Esposito D, Del Vecchio P, Barone G (2001) A thermodynamic study of herring protamine-DNA complex by differential scanning calorimetry. Phys Chem Chem Phys 3: 5320–5325. doi: 10.1039/b107218h
|
| [115] |
Dukhopelnikov EV, Bereznyak EG, Khrebtova AS, et al. (2013) Determination of ligand to DNA binding parameters from two-dimensional DSC curves. J Therm Anal Calorim 111: 1817–1827. doi: 10.1007/s10973-012-2561-6
|
| [116] |
Straume M, Freire E (1992) Two-dimensional differential scanning calorimetry: Simultaneous resolution of intrinsic protein structural energetics and ligand binding interactions by global linkage analysis. Anal Biochem 203: 259–268. doi: 10.1016/0003-2697(92)90311-T
|
| [117] |
Freire E (1994) Statistical thermodynamic analysis of differential scanning calorimetry data: structural deconvolution of heat capacity function of proteins. Methods Enzymol 240: 502–530. doi: 10.1016/S0076-6879(94)40062-8
|
| [118] | Freire E (1995) Differential scanning calorimetry. Methods Mol Biol 40: 191–218. |
| [119] |
Marky LA, Breslauer KJ (1987) Calculating thermodynamic data for transitions of any molecularity from equilibrium melting curves. Biopolymers 26: 1601–1620. doi: 10.1002/bip.360260911
|
| [120] |
Sturtevant JM (1987) Biochemical Applications of Differential Scanning Calorimetry. Annu Rev Phys Chem 38: 463–488. doi: 10.1146/annurev.pc.38.100187.002335
|
| [121] |
Kawai Y (1999) Thermal transition profiles of bacteriophage T4 and its DNA. J Gen Appl Microbiol 45: 135–138. doi: 10.2323/jgam.45.135
|
| [122] |
Tostesen E, Sandve GK, Liu F, et al. (2009) Segmentation of DNA sequences into twostate regions and melting fork regions. J Phys Condens Matter 21: 034109. doi: 10.1088/0953-8984/21/3/034109
|
| [123] | Duguid JG, Bloomfield VA, Benevides JM, et al. (1996) DNA melting investigated by differential scanning calorimetry and Raman spectroscopy. Biophys J 71: 3350–3360. |
| [124] |
Movileanu L, Benevides JM, Thomas GJ (2002) Determination of base and backbone contributions to the thermodynamics of premelting and melting transitions in B DNA. Nucleic Acids Res 30: 3767–3777. doi: 10.1093/nar/gkf471
|
| [125] | Dukhopelnikov EV (2014) Modeling of heat absorption curves for ligand-competitor-DNA triple system. Biophysical Bulletin 31: 49–58. |
| [126] |
1. Finley A, Copeland RA (2014) Small molecule control of chromatin remodeling. Chem Biol 21: 1196-1210. doi: 10.1016/j.chembiol.2014.07.024
|
| [127] |
2. Prinjha R, Tarakhovsky A (2013) Chromatin targeting drugs in cancer and immunity. Genes Dev 27: 1731-1738. doi: 10.1101/gad.221895.113
|
| [128] |
3. Rabbani A, Finn RM, Ausio J (2005) The anthracycline antibiotics: antitumor drugs that alter chromatin structure. Bioessays 27: 50-56. doi: 10.1002/bies.20160
|
| [129] |
4. Majumder P, Pradhan SK, Devi PG, et al. (2007) Chromatin as a target for the DNA-binding anticancer drugs. Subcell Biochem 41: 145-189. doi: 10.1007/1-4020-5466-1_8
|
| [130] |
5. Banerjee A, Majumder P, Sanyal S, et al. (2014) The DNA intercalators ethidium bromide and propidium iodide also bind to core histones. FEBS Open Bio 4: 251-259. doi: 10.1016/j.fob.2014.02.006
|
| [131] |
6. Hajihassan Z, Rabbani-Chadegani A (2009) Studies on the binding affinity of anticancer drug mitoxantrone to chromatin, DNA and histone proteins. J Biomed Sci 16: 31. doi: 10.1186/1423-0127-16-31
|
| [132] |
7. Rabbani-Chadegani A, Chamani E, Hajihassan Z (2009) The effect of vinca alkaloid anticancer drug, vinorelbine, on chromatin and histone proteins in solution. Eur J Pharmacol 613: 34-38. doi: 10.1016/j.ejphar.2009.04.040
|
| [133] |
8. Rabbani-Chadegani A, Keyvani-Ghamsari S, Zarkar N (2011) Spectroscopic studies of dactinomycin and vinorelbine binding to deoxyribonucleic acid and chromatin. Spectrochim Acta A Mol Biomol Spectrosc 84: 62-67. doi: 10.1016/j.saa.2011.08.064
|
| [134] |
9. Rabbani A, Iskandar M, Ausio J (1999) Daunomycin-induced unfolding and aggregation of chromatin. J Biol Chem 274: 18401-18406. doi: 10.1074/jbc.274.26.18401
|
| [135] |
10. Mir MA, Majee S, Das S, et al. (2003) Association of chromatin with anticancer antibiotics, mithramycin and chromomycin A3. Bioorg Med Chem 11: 2791-2801. doi: 10.1016/S0968-0896(03)00211-6
|
| [136] |
11. Mir MA, Dasgupta D (2001) Association of the anticancer antibiotic chromomycin A(3) with the nucleosome: role of core histone tail domains in the binding process. Biochemistry 40: 11578-11585. doi: 10.1021/bi010731r
|
| [137] |
12. Mir MA, Dasgupta D (2001) Interaction of antitumor drug, mithramycin, with chromatin. Biochem Biophys Res Commun 280: 68-74. doi: 10.1006/bbrc.2000.4075
|
| [138] |
13. Hagmar P, Pierrou S, Nielsen P, et al. (1992) Ionic strength dependence of the binding of methylene blue to chromatin and calf thymus DNA. J Biomol Struct Dyn 9: 667-679. doi: 10.1080/07391102.1992.10507947
|
| [139] |
14. Mir MA, Das S, Dasgupta D (2004) N-terminal tail domains of core histones in nucleosome block the access of anticancer drugs, mithramycin and daunomycin, to the nucleosomal DNA. Biophys Chem 109: 121-135. doi: 10.1016/j.bpc.2003.10.023
|
| [140] |
15. Hurley LH (2002) DNA and its associated processes as targets forcancer therapy. Nat Rev Cancer 2: 188-200. doi: 10.1038/nrc749
|
| [141] | 17. Veselkov AN, Maleev VYa, Glibin EN, et al. (2003) Structure-activity relation for synthetic phenoxazone drugs.Evidence for a direct correlation between DNA binding and pro-apoptotic activity. Eur J Biochem 270: 4200-4207 |
| [142] |
18. Andersson J, Lincoln P (2011) Stereoselectivity for DNA threading intercalation of short binuclear ruthenium complexes. J Phys Chem B 115: 14768-14775. doi: 10.1021/jp2062767
|
| [143] | 19. Chaurasiya KR, Paramanathan T, McCauley MJ, et al. (2010) Biophysical characterization of DNA binding from single molecule force measurements. Phys Life Rev 7: 299-341. |
| [144] |
20. Nordell P, Lincoln P (2005) Mechanism of DNA threading intercalation of binuclear Ru complexes: uni- or bimolecular pathways depending on ligand structure and binding density. J Am Chem Soc 127: 9670-9671. doi: 10.1021/ja0521674
|
| [145] | 21. Andersson J, Li M, Lincoln P (2010) AT-specific DNA binding of binuclear ruthenium complexes at the border of threading intercalation. Chemistry 16: 11037-11046. |
| [146] | 22. Wilhelmsson LM, Lincoln P, Nordґen B (2006) Slow DNA binding. In: Waring M, editor. Sequence-Specific DNA Binding Agents, The Royal Society of Chemistry, Cambridge, 69-95. |
| [147] |
23. Palchaudhuri R, Hergenrother PJ (2007) DNA as a target for anticancer compounds: methods to determine the mode of binding and the mechanism of action. Curr Opin Biotechnol 18: 497-503. doi: 10.1016/j.copbio.2007.09.006
|
| [148] | 24. Barton TF, Cooney RP, Denny WA (1992) Surface-enhanced Raman spectroscopic study of amsacrine and amsacrine-DNA interactions. J Raman Spectrosc 23: 341-345. |
| [149] |
25. Rodger A, Blagbrough IS, Adlam G, et al. (1994) DNA binding of a spermine derivative: Spectroscopic study of anthracene-9-carbonyl-n1-spermine with poly[d(G-C)·(d(G-C))] and poly[d(A-T) · d(A-T)]. Biopolymers 34: 1583-1593. doi: 10.1002/bip.360341203
|
| [150] | 26. Hackl EV, Galkin VL, Blagoi YP (2004) DNA interaction with biologically active divalent metal ions: binding constants calculation. Int J Biol Macromol 34: 303-308. |
| [151] |
27. Evstigneev MP, Mykhina YV, Davies DB (2005) Complexation of daunomycin with a DNA oligomer in the presence of an aromatic vitamin (B2) determined by NMR spectroscopy. Biophys Chem 118: 118-127. doi: 10.1016/j.bpc.2005.08.007
|
| [152] |
28. Hackl EV, Blagoi YP (2005) The effect of temperature on DNA structural transitions under the action of Cu2+ and Ca2+ ions in aqueous solutions. Biopolymers 77: 315-324. doi: 10.1002/bip.20225
|
| [153] | 29. Kruglova EB, Gladkovskaia NA, Maleev V (2005) The use of the spectrophotometric analysis for the calculation of the thermodynamic parameters in actinocin derivative-DNA systems. Biofizika 50: 253-264. |
| [154] | 30. Evstigneev MP, Baranovskii SF, Rybakova KA, et al. (2006) 1H NMR study of the complexation of the quinolone antibiotic norfloxacin with DNA. Mol Biol (Mosk) 40: 894-899. |
| [155] |
31. Evstigneev MP, Rybakova KA, Davies DB (2006) Complexation of norfloxacin with DNA in the presence of caffeine. Biophys Chem 121: 84-95. doi: 10.1016/j.bpc.2005.12.003
|
| [156] |
32. Williams AK, Dasilva SC, Bhatta A, et al. (2012) Determination of the drug-DNA binding modes using fluorescence-based assays. Anal Biochem 422: 66-73. doi: 10.1016/j.ab.2011.12.041
|
| [157] |
33. Anupama B, Sunita M, Shiva LD, et al. (2014) Synthesis, spectral characterization, DNA binding studies and antimicrobial activity of Co(II), Ni(II), Zn(II), Fe(III) and VO(IV) complexes with 4-aminoantipyrine Schiff base of ortho-vanillin. J Fluoresc 24: 1067-1076. doi: 10.1007/s10895-014-1386-z
|
| [158] | 34. Zasedatelev AS, Gurskii GV, Vol'kenshtein MV (1971) Theory of one-dimensional adsorption. I. Adsorption of small molecules on a homopolymer. Molecular biology 5: 194-198. |
| [159] |
35. McGhee JD, von Hippel PH (1974) Theoretical aspects of DNA-protein interactions: co-operative and non-co-operative binding of large ligands to a one-dimensional homogeneous lattice. J Mol Biol 86: 469-489. doi: 10.1016/0022-2836(74)90031-X
|
| [160] |
36. Schellman JA (1974) Cooperative Multisite Binding to DNA. Israel Journal of Chemistry 12: 219-238. doi: 10.1002/ijch.197400021
|
| [161] |
37. Nechipurenko YD, Gursky GV (1986) Cooperative effects on binding of proteins to DNA. Biophys Chem 24: 195-209. doi: 10.1016/0301-4622(86)85025-6
|
| [162] |
38. Lando DY, Teif VB (2000) Long-range interactions between ligands bound to a DNA molecule give rise to adsorption with the character of phase transition of the first kind. J Biomol Struct Dyn 17: 903-911. doi: 10.1080/07391102.2000.10506578
|
| [163] |
39. Teif VB, Rippe K (2010) Statistical-mechanical lattice models for protein-DNA binding in chromatin. J Phys Condens Matter 22: 414105. doi: 10.1088/0953-8984/22/41/414105
|
| [164] | 40. Breslauer KJ, Freire E, Straume M (1992) Calorimetry: a tool for DNA and ligand-DNA studies. Methods Enzymol 211: 533-567. |
| [165] |
41. Doyle ML (1997) Characterization of binding interactions by isothermal titration calorimetry. Curr Opin Biotechnol 8: 31-35. doi: 10.1016/S0958-1669(97)80154-1
|
| [166] |
42. Jelesarov I, Bosshard HR (1999) Isothermal titration calorimetry and differential scanning calorimetry as complementary tools to investigate the energetics of biomolecular recognition. J Mol Recognit 12: 3-18. doi: 10.1002/(SICI)1099-1352(199901/02)12:1<3::AID-JMR441>3.0.CO;2-6
|
| [167] | 43. Haq I, Chowdhry BZ, Jenkins TC (2001)Calorimetric techniques in the study of high-order DNA-drug interactions. Methods Enzymol 340: 109-149. |
| [168] | 44. Thomson JA, Ladbury JE (2004) Isothermal titration calorimetry: a tutorial. In Ladbury JE, Doyle ML, editors. Biocalorimetry 2. Applications of Calorimetry in the Biological Sciences, Chichester: John Wiley & Sons, 35-58. |
| [169] | 45. Holdgate GA, Ward WH (2005) Measurements of binding thermodynamics in drug discovery. Drug Discov Today 10: 1543-1550. |
| [170] |
46. Freyer MW, Lewis EA (2008) Isothermal titration calorimetry: experimental design, data analysis, and probing macromolecule/ligand binding and kinetic interactions. Methods Cell Biol 84: 79-113. doi: 10.1016/S0091-679X(07)84004-0
|
| [171] |
47. Bhadra K, Maiti M, Kumar GS (2008) Berberine-DNA complexation: New insights into the cooperative binding and energetic aspects. Biochimica et Biophysica Acta 1780: 1054-1061. doi: 10.1016/j.bbagen.2008.05.005
|
| [172] | 48. Hossain M, Kumar GS (2009) DNA intercalation of methylene blue and quinacrine: new insights into base and sequence specificity from structural and thermodynamic studies with polynucleotides. Mol Biosyst 5: 1311-1322. |
| [173] |
49. Crane-Robinson C, Dragan AI, Read CM (2009) Defining the thermodynamics of protein/DNA complexes and their components using micro-calorimetry. Methods Mol Biol 543: 625-651. doi: 10.1007/978-1-60327-015-1_37
|
| [174] |
50. Kabir A, Kumar GS (2013) Binding of the biogenic polyamines to deoxyribonucleic acids of varying base composition: base specificity and the associated energetics of the interaction. PLoS One 8: e70510. doi: 10.1371/journal.pone.0070510
|
| [175] |
51. Kumar S, Spano MN, Arya DP (2014) Shape readout of AT-rich DNA by carbohydrates. Biopolymers 101: 720-732. doi: 10.1002/bip.22448
|
| [176] |
52. Basu A, Kumar GS (2015) Thermodynamic characterization of proflavine-DNA binding through microcalorimetric studies. J Chem Thermodyn 87: 1-7. doi: 10.1016/j.jct.2015.03.009
|
| [177] | 53. Basu A, Kumar GS (2015) Studies on the interaction of the food colorant tartrazine with double stranded deoxyribonucleic acid. J Biomol Struct Dyn 10: 1-8. |
| [178] | 54. Chaires JB (1997) Possible origin of differences between van't Hoff and calorimetric enthalpy estimates. Biophys Chem 64: 15-23. |
| [179] | 55. Chaires JB (2008) Calorimetry and thermodynamics in drug design. Annu Rev Biophys 37: 135-151. |
| [180] |
56. Janjua NK, Siddiqa A, Yaqub A, et al. (2009) Spectrophotometric analysis of flavonoid-DNA binding interactions at physiological conditions. Spectrochim Acta A Mol Biomol Spectrosc 74: 1135-1137. doi: 10.1016/j.saa.2009.09.022
|
| [181] |
57. Temerk YM, Ibrahim MS, Kotb M (2009) Voltammetric and spectroscopic studies on binding of antitumor Morin, Morin-Cu complex and Morin-beta-cyclodextrin with DNA. Spectrochim Acta A Mol Biomol Spectrosc 71: 1830-1836. doi: 10.1016/j.saa.2008.07.001
|
| [182] |
58. Baranovskii SF, Chernyshev DN, Buchel’nikov AS, et al. (2011) Thermodynamic analysis of complex formation of ethidium bromide with DNA in water solutions. Biophysics 56: 214-219. doi: 10.1134/S0006350911020023
|
| [183] | 59. Chaires JB (1997) Energetics of drug-DNA interactions. Biopolymers 44: 201-215. |
| [184] | 60. Ren J, Jenkins TC, Chaires JB (2000) Energetics of DNA intercalation reactions. Biochemistry 39: 8439-8447. |
| [185] | 61. Davies DB, Veselkov AN (1996) Structural and thermodynamical analysis of molecular complexation by 1H NMR spectroscopy. Intercalation of ethidium bromide with the isomeric deoxytetranucleoside triphosphates 5’-d(GpCpGpC) and 5’-d(CpGpCpG) in aqueous solution. J Chem Soc Faraday Trans 92: 3545-3557. |
| [186] |
62. Kostjukov VV, Pahomov VI, Andrejuk DD, et al. (2007) Investigation of the complexation of the anti-cancer drug novantrone with the hairpin structure of the deoxyheptanucleotide 5′-d(GpCpGpApApGpC). J Mol Struct 843: 78-86. doi: 10.1016/j.molstruc.2006.12.036
|
| [187] |
63. Wartell RM, Benight AS (1985) Thermal denaturation of DNA molecules: A comparison of theory with experiment. Phys Rep 126: 67-107. doi: 10.1016/0370-1573(85)90060-2
|
| [188] |
64. Rice SA, Doty P (1957) The Thermal Denaturation of Desoxyribose Nucleic Acid. J Am Chem Soc 79: 3937-3947. doi: 10.1021/ja01572a001
|
| [189] |
65. Guedin A, Lacroix L, Mergny JL (2010) Thermal melting studies of ligand DNA interactions. Methods Mol Biol 613: 25-35. doi: 10.1007/978-1-60327-418-0_2
|
| [190] | 66. Goldstein G, Stern KG (1950) Experiments on the sonic, thermal, and enzymic depolymerization of desoxyribosenucleic acid. J Struc Chem 5: 687-708. |
| [191] |
67. Thomas R (1954) Recherches sur la d'enaturation des acides desoxyribonucléiques. Biochimica et Biophysica Acta 14: 231-240. doi: 10.1016/0006-3002(54)90163-8
|
| [192] | 68. Frank-Kamenetskii M (1965) Theory of the helix-coil transition for deoxyribonucleic acids with additional connections between the chains. Vysokomolekulyarnye Soedineniya 7: 354-361. |
| [193] | 69. Frank-Kamenetskii M (1968) Consideration of helix-coil transition in homopolymers by the most probable distribution method. Mol Biol 2: 408-419. |
| [194] |
70. Stewart CR (1968) Broadening by acridine orange of the thermal transition of DNA. Biopolymers 6: 1737-1743. doi: 10.1002/bip.1968.360061208
|
| [195] |
71. Lazurkin YS, Frank-Kamenetskii MD, Trifonov EN (1970) Melting of DNA: its study and application as a research method. Biopolymers 9: 1253-1306. doi: 10.1002/bip.1970.360091102
|
| [196] |
72. Barcelo F, Capo D, Portugal J (2002) Thermodynamic characterization of the multivalent binding of chartreusin to DNA. Nucleic Acids Res 30: 4567-4573. doi: 10.1093/nar/gkf558
|
| [197] |
73. Zhong W, Yu JS, Liang Y (2003) Chlorobenzylidine-herring sperm DNA interaction: binding mode and thermodynamic studies. Spectrochim Acta A Mol Biomol Spectrosc 59: 1281-1288. doi: 10.1016/S1386-1425(02)00301-3
|
| [198] | 74. Vardevanyan PO, Antonyan AP, Hambardzumyan LA, et al. (2013) Thermodynamic analysis of DNA complexes with methylene blue, ethidium bromide and Hoechst 33258. Biopolym. Cell 29: 515-520. |
| [199] | 75. Hajian R, Guan Huat T (2013) Spectrophotometric Studies on the Thermodynamics of the ds-DNA Interaction with Irinotecan for a Better Understanding of Anticancer Drug-DNA Interactions. J Spectrosc 2013: 1-8. |
| [200] | 76. Cooper A, Johnson CM (1994) Introduction to microcalorimetry and biomolecular energetics. Methods Mol Biol 22: 109-24. |
| [201] | 77. Rosgen J, Hinz HJ (1999) Theory and practice of DSC mesuarements on proteins. In: Kemp RB, editor. Handbook of Thermal Analysis and Calorimetry, Vol.4, From Macromolecules to Man, Amsterdam: Elsevier, 63-108. |
| [202] |
78. Bruylants G, Wouters J, Michaux C (2005) Differential scanning calorimetry in life science: thermodynamics, stability, molecular recognition and application in drug design. Curr Med Chem 12: 2011-. doi: 10.2174/0929867054546564
|
| [203] |
79. Spink CH (2008) Differential scanning calorimetry. Methods Cell Biol 84: 115-141. doi: 10.1016/S0091-679X(07)84005-2
|
| [204] |
80. Bereznyak EG, Gladkovskaya NA, Khrebtova AS, et al. (2009) Peculiarities of DNA-proflavine binding under different concentration ratios. Biophysics 54: 574-580. doi: 10.1134/S0006350909050030
|
| [205] | 81. Garbett N (2011) The Use of Calorimetry to Study Ligand-DNA Interactions. In: Aldrich-Wright J, editor. Metallointercalators, Vienna: Springer, 299-324. |
| [206] |
82. Ising E (1925) Beitrag zur Theorie des Ferromagnetismus. Zeitschrift für Physik 31: 253-258. doi: 10.1007/BF02980577
|
| [207] | 83. Poland DC, Scheraga HA (1970) The theory of helix coil transition, New York: Academic Press. |
| [208] | 84. Peyrard M, Bishop AR (1989) Statistical mechanics of a nonlinear model for DNA denaturation. Phys Rev Lett 62: 2755-2758. |
| [209] | 85. Dauxois T, Peyrard M, Bishop AR (1993) Entropy-driven DNA denaturation. Physical Review E 47: R44-R47. |
| [210] | 86. Dauxois T, Peyrard M, Bishop AR (1993) Dynamics and thermodynamics of a nonlinear model for DNA denaturation. Phys Rev E Stat Phys Plasmas Fluids Relat Interdiscip Topics 47: 684-695. |
| [211] | 87. Grosberg AIU, Khokhlov AR (1994) Statistical physics of macromolecules, New York: AIP Press. |
| [212] | 88. Frank-Kamenetskii MD, Prakash S (2014) Fluctuations in the DNA double helix: A critical review. Phys Life Rev 11: 153-170. |
| [213] | 89. Cantor CR, Schimmel PR (1980) Biophysical chemistry. 3. The behavior of biological macromolecules, San Francisco, Freeman. |
| [214] | 90. Zimm BH, Bragg JK (1959) Theory of the Phase Transition between Helix and Random Coil in Polypeptide Chains. J Chem Phys 31: 526-535. |
| [215] | 91. Crothers DM (1971) Statistical thermodynamics of nucleic acid melting transitions with coupled binding equilibria. Biopolymers 10: 2147-2160. |
| [216] | 92. McGhee JD (1976) Theoretical calculations of the helix-coil transition of DNA in the presence of large, cooperatively binding ligands. Biopolymers 15: 1345-1375. |
| [217] | 93. Akhrem AA, Fridman AS, Lando D (1985) Theory of helix-coil transition of the heterogeneous DNA-heteroqeneous ligands complexes. Biopolym Cell 1: 171-179. |
| [218] |
94. Lando D (1994) A theoretical consideration of the influence of selective binding of small ligands on DNA helix-coil transition. J Biomol Struct Dyn 12: 343-354. doi: 10.1080/07391102.1994.10508744
|
| [219] | 95. Akhrem AA, Lando D (1979) Influence of ligands characteristic of selective binding to a certain type of base pairs on DNA helix-coil transition I. Model. Theory. Mol Biol (Mosk) 13: 1098-1109. |
| [220] | 96. Akhrem AA, Lando D, Shpakovskii AG, et al. (1990) The effect of long-range interactions between adsorbed ligands on the DNA helix-coil transition. Mol Biol (Mosk) 24: 649-656. |
| [221] | 97. Lando D, Ivanova MA, Akhrem AA (1980) Effect of changes in the stoichiometry of DNA-ligand complexes during heat denaturation of DNA on helix-coil transition parameters. Mol Biol (Mosk) 14: 1281-1288. |
| [222] |
98. Karapetian AT, Mehrabian NM, Terzikian GA, et al. (1996) Theoretical treatment of melting of complexes of DNA with ligands having several types of binding sites on helical and single-stranded DNA. J Biomol Struct Dyn 14: 275-283. doi: 10.1080/07391102.1996.10508118
|
| [223] |
99. Plum GE, Bloomfield VA (1990) Structural and electrostatic effects on binding of trivalent cations to double-stranded and single-stranded poly[d (AT)]. Biopolymers 29: 13-27. doi: 10.1002/bip.360290105
|
| [224] |
100. Spink CH, Chaires JB (1997) Thermodynamics of the Binding of a Cationic Lipid to DNA. J Am Chem Soc 119: 10920-10928. doi: 10.1021/ja964324s
|
| [225] |
101. Leng F, Chaires JB, Waring MJ (2003) Energetics of echinomycin binding to DNA. Nucleic Acids Res 31: 6191-6197. doi: 10.1093/nar/gkg826
|
| [226] |
102. Pasic L, Sepcic K, Turk T, et al. (2001) Characterization of parazoanthoxanthin A binding to a series of natural and synthetic host DNA duplexes. Arch Biochem Biophys 393: 132-142. doi: 10.1006/abbi.2001.2469
|
| [227] |
103. Portugal J, Cashman DJ, Trent JO, et al. (2005) A new bisintercalating anthracycline with picomolar DNA binding affinity. J Med Chem 48: 8209-8219. doi: 10.1021/jm050902g
|
| [228] |
104. Liu Y-J, Wei X, Mei W-J, et al. (2007) Synthesis, characterization and DNA binding studies of ruthenium(II) complexes: [Ru(bpy)2(dtmi)]2+ and [Ru(bpy)2(dtni)]2+. Transit Metal Chem 32: 762-768. doi: 10.1007/s11243-007-0246-y
|
| [229] |
105. Peng B, Chen X, Du KJ, et al. (2009) Synthesis, characterization and DNA-binding studies of ruthenium(II) mixed-ligand complexes containing dipyrido[1,2,5]oxadiazolo[3,4-b]quinoxaline. Spectrochim Acta A Mol Biomol Spectrosc 74: 896-901. doi: 10.1016/j.saa.2009.08.031
|
| [230] | 106. Barcelo F, Portugal J (2004) Elsamicin A binding to DNA. A comparative thermodynamic characterization. FEBS Lett 576: 68-72. |
| [231] |
107. Barcelo F, Scotta C, Ortiz-Lombardia M, et al. (2007) Entropically-driven binding of mithramycin in the minor groove of C/G-rich DNA sequences. Nucleic Acids Res 35: 2215-2226. doi: 10.1093/nar/gkm037
|
| [232] | 108. Marky LA, Blumenfeld KS, Breslauer KJ (1983) Calorimetric and spectroscopic investigation of drug-DNA interactions. I. The binding of netropsin to poly d(AT). Nucleic Acids Res 11: 2857-2870. |
| [233] |
109. Remeta DP, Mudd CP, Berger RL, et al (1993) Thermodynamic characterization of daunomycin-DNA interactions: comparison of complete binding profiles for a series of DNA host duplexes. Biochemistry 32: 5064-5073. doi: 10.1021/bi00070a014
|
| [234] | 110. Brandts JF, Lin LN (1990) Study of strong to ultratight protein interactions using differential scanning calorimetry. Biochemistry 29: 6927-6940. |
| [235] | 111. Barone G, Catanzano F, Del Vecchio P, et al. (1995) Differential scanning calorimetry as a tool to study protein-ligand interactions. Pure Appl Chem 67: 1867-1872. |
| [236] |
112. Dassie SA, Celej MS, Fidelio GD (2005) Protein Unfolding Coupled to Ligand Binding: Differential Scanning Calorimetry Simulation Approach. J Chem Educ 82: 85. doi: 10.1021/ed082p85
|
| [237] |
113. Celej MS, Dassie SA, Gonzalez M, et al. (2006) Differential scanning calorimetry as a tool to estimate binding parameters in multiligand binding proteins. Anal Biochem 350: 277-284. doi: 10.1016/j.ab.2005.12.029
|
| [238] |
114. Esposito D, Del Vecchio P, Barone G (2001) A thermodynamic study of herring protamine-DNA complex by differential scanning calorimetry. Phys Chem Chem Phys 3: 5320-5325. doi: 10.1039/b107218h
|
| [239] |
115. Dukhopelnikov EV, Bereznyak EG, Khrebtova AS, et al. (2013) Determination of ligand to DNA binding parameters from two-dimensional DSC curves. J Therm Anal Calorim 111: 1817-1827. doi: 10.1007/s10973-012-2561-6
|
| [240] |
116. Straume M, Freire E (1992) Two-dimensional differential scanning calorimetry: Simultaneous resolution of intrinsic protein structural energetics and ligand binding interactions by global linkage analysis. Anal Biochem 203: 259-268. doi: 10.1016/0003-2697(92)90311-T
|
| [241] |
117. Freire E (1994) Statistical thermodynamic analysis of differential scanning calorimetry data: structural deconvolution of heat capacity function of proteins. Methods Enzymol 240: 502-530. doi: 10.1016/S0076-6879(94)40062-8
|
| [242] | 118. Freire E (1995) Differential scanning calorimetry. Methods Mol Biol 40: 191-218. |
| [243] |
119. Marky LA, Breslauer KJ (1987) Calculating thermodynamic data for transitions of any molecularity from equilibrium melting curves. Biopolymers 26: 1601-1620. doi: 10.1002/bip.360260911
|
| [244] |
120. Sturtevant JM (1987) Biochemical Applications of Differential Scanning Calorimetry. Annu Rev Phys Chem 38: 463-488. doi: 10.1146/annurev.pc.38.100187.002335
|
| [245] |
121. Kawai Y (1999) Thermal transition profiles of bacteriophage T4 and its DNA. J Gen Appl Microbiol 45: 135-138. doi: 10.2323/jgam.45.135
|
| [246] |
122. Tostesen E, Sandve GK, Liu F, et al. (2009) Segmentation of DNA sequences into twostate regions and melting fork regions. J Phys Condens Matter 21: 034109. doi: 10.1088/0953-8984/21/3/034109
|
| [247] | 123. Duguid JG, Bloomfield VA, Benevides JM, et al. (1996) DNA melting investigated by differential scanning calorimetry and Raman spectroscopy. Biophys J 71: 3350-3360. |
| [248] |
124. Movileanu L, Benevides JM, Thomas GJ (2002) Determination of base and backbone contributions to the thermodynamics of premelting and melting transitions in B DNA. Nucleic Acids Res 30: 3767-3777. doi: 10.1093/nar/gkf471
|
| [249] | 125. Dukhopelnikov EV (2014) Modeling of heat absorption curves for ligand-competitor-DNA triple system. Biophysical Bulletin 31: 49-58. |
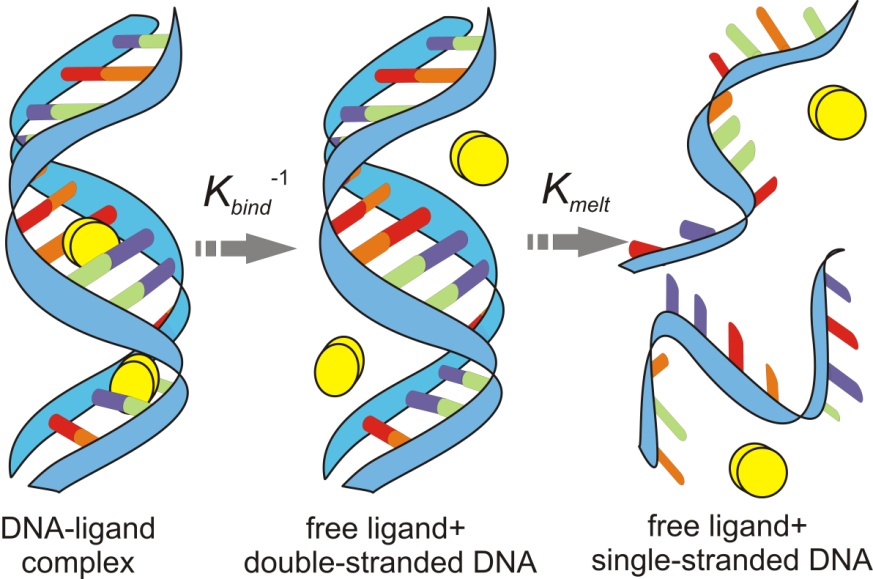
Figures(1)

Ekaterina Bereznyak, Natalia Gladkovskaya, Evgeniy Dukhopelnykov, Anastasiya Gerus, Anastasiya Lantushenko, Maxim Evstigneev. Thermal analysis of ligand-DNA interaction: determination of binding parameters[J]. AIMS Biophysics, 2015, 2(4): 423-440. doi: 10.3934/biophy.2015.4.423







 DownLoad:
DownLoad: