Citation: Chiara Ceresa, Francesco Tessarolo, Devid Maniglio, Iole Caola, Giandomenico Nollo, Maurizio Rinaldi, Letizia Fracchia. Inhibition of Candida albicans biofilm by lipopeptide AC7 coated medical-grade silicone in combination with farnesol[J]. AIMS Bioengineering, 2018, 5(3): 192-208. doi: 10.3934/bioeng.2018.3.192

| [1] |
Liu Y, Filler SG (2011) Candida albicans Als3, a multifunctional adhesin and invasin. Eukaryot Cell 10: 168-173. doi: 10.1128/EC.00279-10

|
| [2] | Hawser SP, Douglas LJ (1994) Biofilm formation by Candida species on the surface of catheter materials in vitro. Infect Immun 62: 915-921. |
| [3] |
Ramage G, Martínez JP, López-Ribot JL (2006) Candida biofilms on implanted biomaterials: A clinically significant problem. FEMS Yeast Res 6: 979-986. doi: 10.1111/j.1567-1364.2006.00117.x
|
| [4] |
Chandra J, Mukherjee PK, Ghannoum MA (2008) In vitro growth and analysis of Candida biofilms. Nat Protoc 3: 1909-1924. doi: 10.1038/nprot.2008.192
|
| [5] |
Finkel JS, Mitchell AP (2011) Genetic control of Candida albicans biofilm development. Nat Rev Microbiol 9: 109-118. doi: 10.1038/nrmicro2475
|
| [6] |
Chandra J, Kuhn DM, Mukherjee PK, et al. (2001) Biofilm formation by the fungal pathogen Candida albicans: Development, architecture, and drug resistance. J Bacteriol 183: 5385-5394. doi: 10.1128/JB.183.18.5385-5394.2001
|
| [7] |
Lazzell AL, Chaturvedi AK, Pierce CG, et al. (2009) Treatment and prevention of Candida albicans biofilms with caspofungin in a novel central venous catheter murine model of candidiasis. J Antimicrob Chemother 64: 567-570. doi: 10.1093/jac/dkp242
|
| [8] |
Francolini I, Donelli G (2010) Prevention and control of biofilm-based medical-device-related infections. FEMS Immunol Med Microbiol 59: 227-238. doi: 10.1111/j.1574-695X.2010.00665.x
|
| [9] | Muakkassa FK, Ghannoum M, (2016) Updates on Therapeutic Strategies Against Candida (and Aspergillus) Biofilm Related Infections, In: Imbert C, editor, Fungal Biofilms and related infections. Advances in Experimental Medicine and Biology, Cham: Springer, 95-103. |
| [10] |
Giles C, Lamont-Friedrich SJ, Michl TD, et al. (2018) The importance of fungal pathogens and antifungal coatings in medical device infections. Biotechnol Adv 36: 264-280. doi: 10.1016/j.biotechadv.2017.11.010
|
| [11] |
Ramage G, Saville SP, Wickes BL, et al. (2002) Inhibition of Candida albicans biofilm formation by farnesol, a quorum-sensing molecule. Appl Environ Microbiol 68: 5459-5463. doi: 10.1128/AEM.68.11.5459-5463.2002
|
| [12] |
Hornby JM, Jensen EC, Lisec AD, et al. (2001) Quorum sensing in the dimorphic fungus Candida albicans is mediated by farnesol. Appl Environ Microbiol 67: 2982-2992. doi: 10.1128/AEM.67.7.2982-2992.2001
|
| [13] |
Nickerson KW, Atkin AL, Hornby JM (2006) Quorum sensing in dimorphic fungi: Farnesol and beyond. Appl Environ Microbiol 72: 3805-3813. doi: 10.1128/AEM.02765-05
|
| [14] |
Deveau A, Hogan DA (2011) Linking quorum sensing regulation and biofilm formation by Candida albicans. Methods Mol Biol 692: 219-233. doi: 10.1007/978-1-60761-971-0_16
|
| [15] |
Donadio S, Monciardini P, Alduina R, et al. (2002) Microbial technologies for the discovery of novel bioactive metabolites. J Biotechnol 99: 187-198. doi: 10.1016/S0168-1656(02)00209-2
|
| [16] |
Singh P, Cameotra SS (2004) Potential applications of microbial surfactants in biomedical sciences. Trends Biotechnol 22: 142-146. doi: 10.1016/j.tibtech.2004.01.010
|
| [17] |
Bĕhal V (2006) Mode of action of microbial bioactive metabolites. Folia Microbiol 51: 359-369. doi: 10.1007/BF02931577
|
| [18] |
Rodrigues L, Van dMH, Teixeira J, et al. (2004) Biosurfactant from Lactococcus lactis 53 inhibits microbial adhesion on silicone rubber. Appl Microbiol Biotechnol 66: 306-311. doi: 10.1007/s00253-004-1674-7
|
| [19] |
Gudiña EJ, Rocha V, Teixeira JA (2010) Isolation and functional characterization of a biosurfactant produced by Lactobacillus paracasei. Colloids Surf B 76: 298-304. doi: 10.1016/j.colsurfb.2009.11.008
|
| [20] |
Janek T, Łukaszewicz M, Krasowska A (2012) Anti-adhesive activity of the biosurfactant pseudofactin II secreted by the Arctic bacterium Pseudomonas fluorescens BD5. BMC Microbiol 12: 24. doi: 10.1186/1471-2180-12-24
|
| [21] | Martinotti MG, Allegrone G, Cavallo M, et al. (2013) Biosurfactants, In: Piemonte V, De Falco M, Basile A, editors, Sustainable Development in Chemical Engineering-Innovative Technologies, Chichester: John Wiley & Sons, 199-240. |
| [22] |
Fracchia L, Banat JJ, Cavallo M, et al. (2015) Potential therapeutic applications of microbial surface-active compounds. AIMS Bioeng 2: 144-162. doi: 10.3934/bioeng.2015.3.144
|
| [23] |
Banat IM, Makkar RS, Cameotra SS (2000) Potential commercial applications of microbial surfactants. Appl Microbiol Biotechnol 53: 495-508. doi: 10.1007/s002530051648
|
| [24] |
Banat IM, Franzetti A, Gandolfi I, et al. (2010) Microbial biosurfactants production, applications and future potential. Appl Microbiol Biotechnol 87: 427-444. doi: 10.1007/s00253-010-2589-0
|
| [25] | Fracchia L, Cavallo M, Martinotti MG, et al. (2012) Biosurfactants and bioemulsifiers: Biomedical and related applications-present status and future potentials, In: Ghista DN, editor, Biomedical Science, Engineering and Technology, Croatia, Rijeka: InTech. |
| [26] | Fracchia L, Ceresa C, Franzetti A, et al. (2014) Industrial Applications of Biosurfactants, In: Kosaric N, Sukan FV, editors, BIOSURFACTANTS. Production and Utilization-Processes, Technologies, and Economics, USA: CRS Press-Taylor & Francis Group, 245-267. |
| [27] |
Sotirova AV, Spasova DI, Galabova DN, et al. (2008) Rhamnolipid-biosurfactant permeabilizing effects on gram-positive and gram-negative bacterial strains. Curr Microbiol 56: 639-644. doi: 10.1007/s00284-008-9139-3
|
| [28] |
Ortiz A, Teruel JA, Espuny MJ, et al. (2009) Interactions of a bacterial biosurfactant trehalose lipid with phosphatidylserine membranes. Chem Phys Lipids 158: 46-53. doi: 10.1016/j.chemphyslip.2008.11.001
|
| [29] |
Zaragoza A, Aranda FJ, Espuny MJ, et al. (2009) A mechanism of membrane permeabilization by a bacterial trehalose lipid biosurfactant produced by Rhodococcus sp. Langmuir 25: 7892-7898. doi: 10.1021/la900480q
|
| [30] |
Sánchez M, Aranda FJ, Teruel JA, et al. (2010) Permeabilization of biological and artificial membranes by a bacterial dirhamnolipid produced by Pseudomonas aeruginosa. J Colloid Interface Sci 341: 240-247. doi: 10.1016/j.jcis.2009.09.042
|
| [31] |
Rodrigues L, Banat IM, Teixeira J, et al. (2006) Biosurfactants: Potential applications in medicine. J Antimicrob Chemother 57: 609-618. doi: 10.1093/jac/dkl024
|
| [32] |
Banat IM, Rienzo MAD, Quinn GA (2014) Microbial biofilms, biosurfactants as antibiofilm agents. Appl Microbiol Biotechnol 98: 9915-9929. doi: 10.1007/s00253-014-6169-6
|
| [33] |
Ceresa C, Rinaldi M, Chiono V, et al. (2016) Lipopeptides from Bacillus subtilis AC7 inhibit adhesion and biofilm formation of Candida albicans on silicone. Antonie Van Leeuwenhoek 109: 1375-1388. doi: 10.1007/s10482-016-0736-z
|
| [34] | Rivardo F, Turner RJ, Allegrone G, et al. (2009) Anti-adhesion activity of two biosurfactants produced by Bacillus spp. prevents biofilm formation of human bacterial pathogens. Appl Microbiol Biotechnol 83: 541-553. |
| [35] |
Ceresa C, Tessarolo F, Caola I, et al. (2015) Inhibition of Candida albicans adhesion on medical-grade silicone by a Lactobacillus-derived biosurfactant. J Appl Microbiol 118: 1116-1125. doi: 10.1111/jam.12760
|
| [36] |
Comoglio F, Fracchia L, Rinaldi M (2013) Bayesian inference from count data using discrete uniform priors. PLoS One 8: e74388. doi: 10.1371/journal.pone.0074388
|
| [37] |
Quinn GA, Maloy AP, Banat MM, et al. (2013) A comparison of effects of broad-spectrum antibiotics and biosurfactants on established bacterial biofilms. Curr Microbiol 67: 614-623. doi: 10.1007/s00284-013-0412-8
|
| [38] |
Rodrigues L, Banat IM, Teixeira J, et al. (2007) Strategies for the prevention of microbial biofilm formation on silicone rubber voice prostheses. J Biomed Mater Res B Appl Biomater 81B: 358-370. doi: 10.1002/jbm.b.30673
|
| [39] |
Ceresa C, Rinaldi M, Fracchia L (2017) Synergistic activity of antifungal drugs and lipopeptide AC7 against Candida albicans biofilm on silicone. AIMS Bioeng 4: 318-334. doi: 10.3934/bioeng.2017.2.318
|
| [40] |
Jabra-Rizk MA, Shirtliff M, James C, et al. (2006) Effect of farnesol on Candida dubliniensis biofilm formation and fluconazole resistance. FEMS Yeast Res 6: 1063-1073. doi: 10.1111/j.1567-1364.2006.00121.x
|
| [41] |
Lara HH, Romero-Urbina DG, Pierce C, et al. (2015) Effect of silver nanoparticles on Candida albicans biofilms: An ultrastructural study. J Nanobiotechnol 13: 91. doi: 10.1186/s12951-015-0147-8
|
| [42] |
Nieminen MT, Novak-Frazer L, Rautemaa V, et al. (2014) A novel antifungal is active against Candida albicans biofilms and inhibits mutagenic acetaldehyde production in vitro. PLoS One 9: e97864. doi: 10.1371/journal.pone.0097864
|
| [43] |
Cochis A, Fracchia L, Martinotti MG, et al. (2012) Biosurfactants prevent in vitro Candida albicans biofilm formation on resins and silicon materials for prosthetic devices. Oral Surg Oral Med Oral Pathol Oral Radiol 113: 755-761. doi: 10.1016/j.oooo.2011.11.004
|
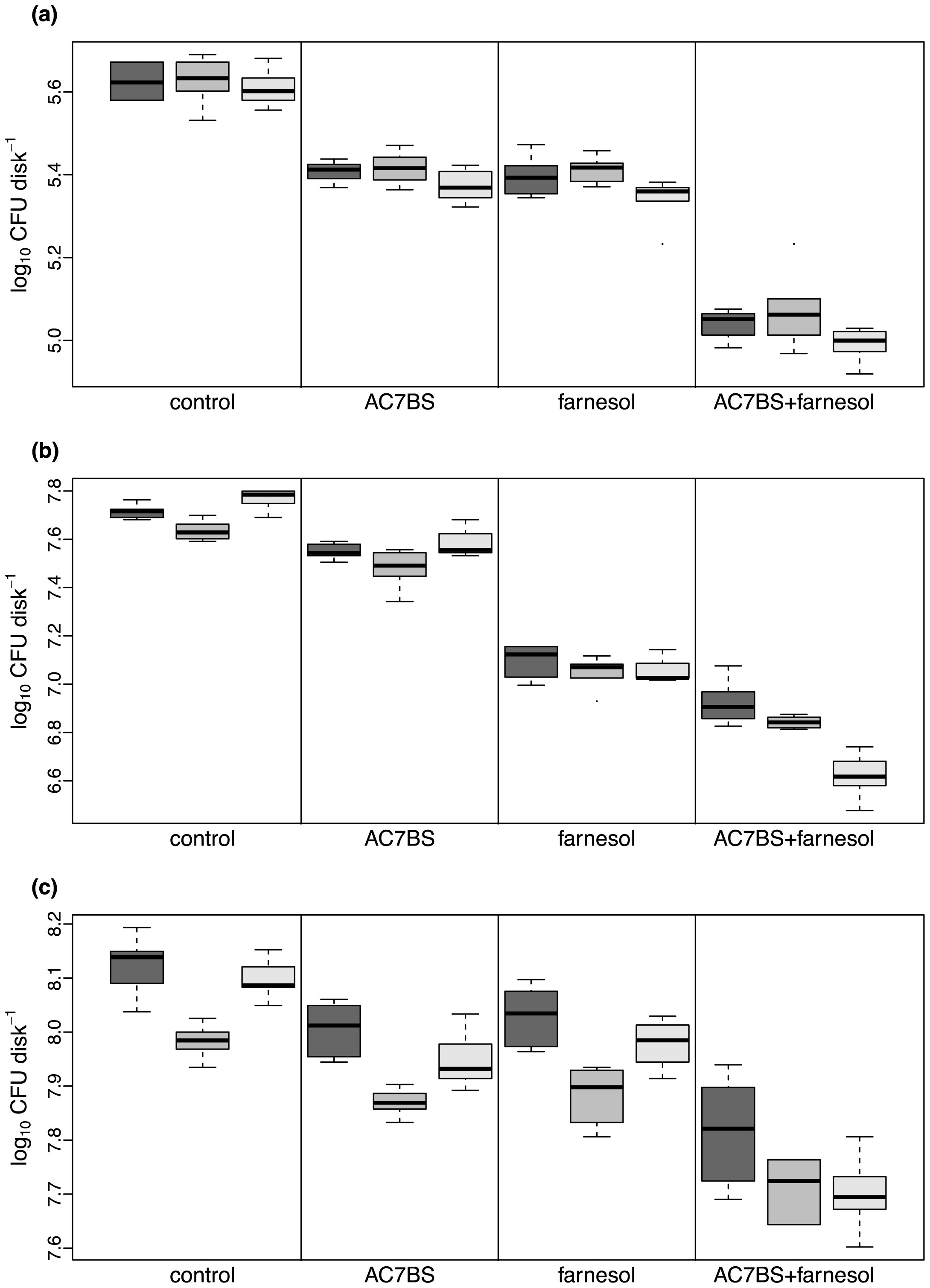
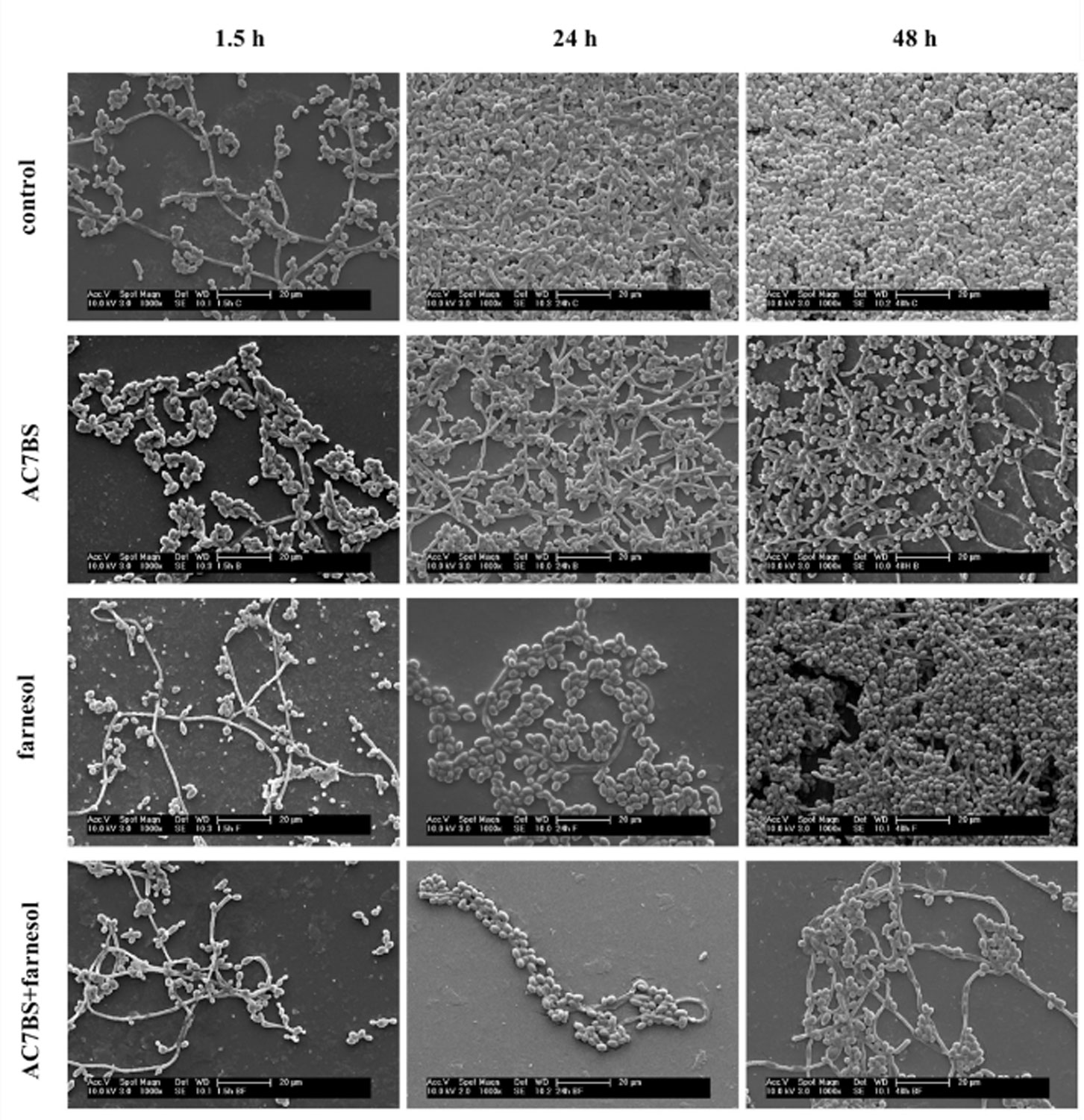
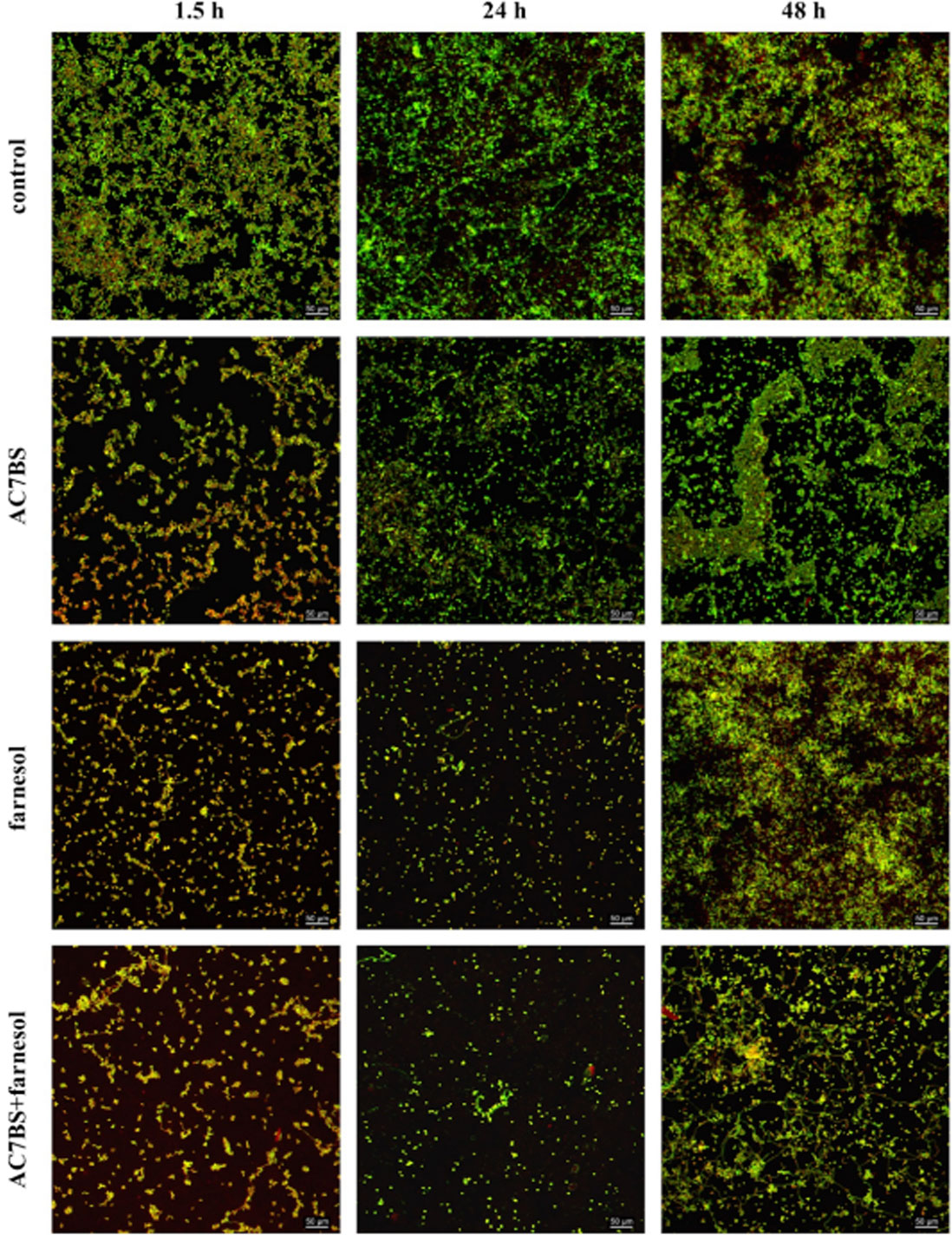

Figures(4) / Tables(2)

Chiara Ceresa, Francesco Tessarolo, Devid Maniglio, Iole Caola, Giandomenico Nollo, Maurizio Rinaldi, Letizia Fracchia. Inhibition of Candida albicans biofilm by lipopeptide AC7 coated medical-grade silicone in combination with farnesol[J]. AIMS Bioengineering, 2018, 5(3): 192-208. doi: 10.3934/bioeng.2018.3.192







 DownLoad:
DownLoad: