Citation: Mohamed Neifar, Rim Chatter, Habib Chouchane, Raya Genouiz, Atef Jaouani, Ahmed Slaheddine Masmoudi, Ameur Cherif. Optimization of enzymatic saccharification of Chaetomorpha linum biomass for the production of macroalgae-based third generation bioethanol[J]. AIMS Bioengineering, 2016, 3(3): 400-411. doi: 10.3934/bioeng.2016.3.400

| [1] |
Demirbas MF (2011) Biofuels from Algae for sustainable Development. Appl Energ 88: 3473–3480. doi: 10.1016/j.apenergy.2011.01.059

|
| [2] |
Milledge JJ, Smith B, Dyer PW, et al. (2014) Macroalgae-Derived Biofuel: A Review of Methods of Energy Extraction from Seaweed Biomass. Energies 7: 7194–7222. doi: 10.3390/en7117194
|
| [3] | Tsuji A, Tominaga K, Nishiyama N, et al. (2013) Comprehensive Enzymatic Analysis of the Cellulolytic System in Digestive Fluid of the Sea Hare Aplysia kurodai. Efficient Glucose Release from Sea Lettuce by Synergistic Action of 45 kDa Endoglucanase and 210 kDa ?-Glucosidase. PLoS ONE 8: e65418. |
| [4] |
Meinita MDN, Kang JY, Jeong GT, et al. (2012) Bioethanol production from the acid hydrolysate of the carrageenophyte Kappaphycus alvarezii (cottonii). J App Phy 24: 857–862. doi: 10.1007/s10811-011-9705-0
|
| [5] |
Borines MG, Leon RL, Cuello JL (2013) Bioethanol production from the macroalgae Sargassum spp. Bioresource Technol 138: 22–29. doi: 10.1016/j.biortech.2013.03.108
|
| [6] |
Khambhaty Y, Mody K, Gandhi MR, et al. (2012) Kappaphycus alvarezii as a source of bioethanol. Bioresource Technol 103: 180–185. doi: 10.1016/j.biortech.2011.10.015
|
| [7] | Kim HM, Wi SG, Jung S, et al. (2015) Efficient approach for bioethanol production from red seaweed Gelidium amansii. Bioresource Technol 175: 128–134. |
| [8] |
Cho YK, Kim H, Kim SK (2013) Bioethanol production from brown seaweed, Undaria pinnatifida, using NaCl acclimated yeast. Bioproc Biosyst Eng 36: 713–719. doi: 10.1007/s00449-013-0895-5
|
| [9] |
Lee JY, Kim YS, Um BH, et al. (2013) Pretreatment of Laminaria japonica for bioethanol production with extremely low acid concentration. Renew Energ 54: 196–200. doi: 10.1016/j.renene.2012.08.025
|
| [10] |
Li K, Liu S, Liu X (2014) An overview of algae bioethanol production. Int J Energ Res 38: 965–977. doi: 10.1002/er.3164
|
| [11] |
Trivedi N, Gupta V, Reddy CRK, et al. (2013) Enzymatic hydrolysis and production of bioethanol from common macrophytic green alga Ulva fasciata Delile. Bioresource Technol 150: 106–112. doi: 10.1016/j.biortech.2013.09.103
|
| [12] |
Schultz-Jensen N, Thygesen A, Leipold F, et al. (2013) Pretreatment of the macroalgae Chaetomorpha linum for the production of bioethanol-Comparison of five pretreatment technologies. Bioresource Technol 140: 36–42. doi: 10.1016/j.biortech.2013.04.060
|
| [13] |
Milledge JJ, Harvey PJ (2016) Potential process ‘hurdles’ in the use of macroalgae as feedstock for biofuel production in the British Isles. J Chem Technol Biot 91:2221–2234. doi: 10.1002/jctb.5003
|
| [14] |
Yanagisawa M, Kawai S, Murata K (2013) Strategies for the production of high concentrations of bioethanol from seaweeds. Bioengineered 4: 224–235. doi: 10.4161/bioe.23396
|
| [15] |
Jang JS, Cho YK, Jeong GT, et al. (2012) Optimization of saccharification and ethanol production by simultaneous saccharification and fermentation (SSF) from seaweed Saccharina japonica. Bioproc Biosyst Eng 35: 11–18. doi: 10.1007/s00449-011-0611-2
|
| [16] | Myers RH, Montgomery DC, Anderson-Cook CM (2009) Response Surface Methodology: Process and Product Optimization Using Designed Experiments, Wiley, New York, USA. |
| [17] | Myers RH, Montgomery DC (1995) Response Surface Methodology: Process and Product Optimization Using Designed Experiments, Wiley, New York, USA. |
| [18] | Mathieu, Phan-Tan-Luu R (1997) Approche méthodologique des surfaces de réponse. In Plans d’expériences: Application à l’entreprise, Technip, Paris, France, 211–278. |
| [19] | Goupy J (1999) Plans d’Expériences Pour Surfaces de Réponse, Dunod, Paris, France. |
| [20] | Tye YY, Lee KT, Abdullah WNW, et al. (2015) Effects of process parameters of various pretreatments on enzymatic hydrolysability of Ceiba pentandra (L.) Gaertn. (Kapok) fibre: A response surface methodology study. Biomass Bioenerg 75: 301–313. |
| [21] | Ra CH, Kim SK (2013) Optimization of pretreatment conditions and use of a two-stage fermentation process for the production of ethanol from seaweed, Saccharina japonica. Biotech Bioproc Eng 18: 715–720. |
| [22] | AOAC International (2005) Official methods of analysis of AOAC International, 18th edn., Revision I, Gaithersburg, MD. |
| [23] |
Dubois M, Gilles KA, Hamilton JK, et al. (1956) Colorimetric Method for Determination of Sugars and Related Substances. Anal Chem 28: 350–356. doi: 10.1021/ac60111a017
|
| [24] | Angell AR, Mata L, Nys RD, et al. (2016) The protein content of seaweeds: a universal nitrogen-to-protein conversion factor of five. J App Phycol 28: 511–524. |
| [25] | Prosky L, Asp NG, Schweizer TF, et al. (1988) Determination of insoluble, soluble, and total dietary fiber in foods and food products: interlaboratory study. J Assoc Anal Chem 71: 1017–1023. |
| [26] |
Chirapart A, Praiboon J, Puangsombat P, et al. (2014) Chemical composition and ethanol production potential of Thai seaweed species. J Appl Phycol 26: 979–986. doi: 10.1007/s10811-013-0235-9
|
| [27] | Miller GL (1959) Use of dinitrosalicylic acid reagent for determination of reducing sugars. Anal Chem 31: 426–428. |
| [28] | Idrees M, Adnan A, Sheikh S, et al. (2013) Optimization of dilute acid pretreatment of water Hyacinth biomass for enzymatic hydrolysis and ethanol production. J Excli 12: 30–40. |
| [29] | Neifar M, Jaouani A, Kamoun A, et al. (2011) Decolorization of Solophenyl red 3BL polyazo dye by laccase-mediator system: Optimization through response surface methodology. Enzyme Res doi:10.4061/2011/179050. |
| [30] | Neifar M, Kamoun A, Jaouani A, et al. (2011) Application of Asymetrical and Hoke designs for optimization of laccase production by the white-rot fungus Fomes fomentarius in solid-state fermentation. Enzyme Res doi:10.4061/2011/368525. |
| [31] | Mathieu D, Nony J, Phan-Tan-Luu R (2000) NEMROD-W Software, LPRAI, Marseille, France. |
| [32] |
Yahmed NB, Jmel MA, Alaya MB, et al. (2016) A biorefinery concept using the green macroalgae Chaetomorpha linum for the coproduction of bioethanol and biogas. Ener Convers Manage 119: 257–265. doi: 10.1016/j.enconman.2016.04.046
|
| [33] | Neifar M, Chouchane H, Maktouf S, et al. (2016) Improved sugar yield for bioethanol production by modelling enzymatic hydrolysis of Peganum Harmala biomass through response surface methodology. Ijes 5: 22–28. |
| [34] |
Wang X, Liu X, Wang G (2011) Two-stage hydrolysis of invasive algal feedstock for ethanol fermentation. J Integr Plant Biol 53: 246–252. doi: 10.1111/j.1744-7909.2010.01024.x
|
| [35] |
Lee JY, Li P, Lee J, et al. (2013) Ethanol production from Saccharina japonica using an extremely low acid pretreatment followed by simultaneous saccharification and fermentation. Bioresource Technol 127: 119–125. doi: 10.1016/j.biortech.2012.09.122
|
| [36] | Coppalo F, Simoncini E, Pulselli RM (2009) Bioethanol potentials from marine residual biomass: an energy evaluation, In: Brebbia, Tiezzi, Eds., Fifth International Conference on Ecosystems and Sustainable Development, Italy. |
| [37] | Majumder A, Singh A, Goyal A (2009) Application of response surface methodology for glucan production from Leuconostoc dextranicum and its structural characterization. Carbohyd Polym 75: 150–156. |
| [38] |
Ferreira SLC, Bruns RE, Ferreira HS, et al. (2007) Box-Behnken design: An alternative for the optimization of analytical methods. Anal Chim Acta 597: 179–186. doi: 10.1016/j.aca.2007.07.011
|
| [39] |
Okuda K, Oka K, Onda A, et al. (2008) Hydrothermal fractional pretreatment of sea algae and its enhanced enzymatic hydrolysis. Chem J Technol Biotechnol 83: 836–41. doi: 10.1002/jctb.1877
|
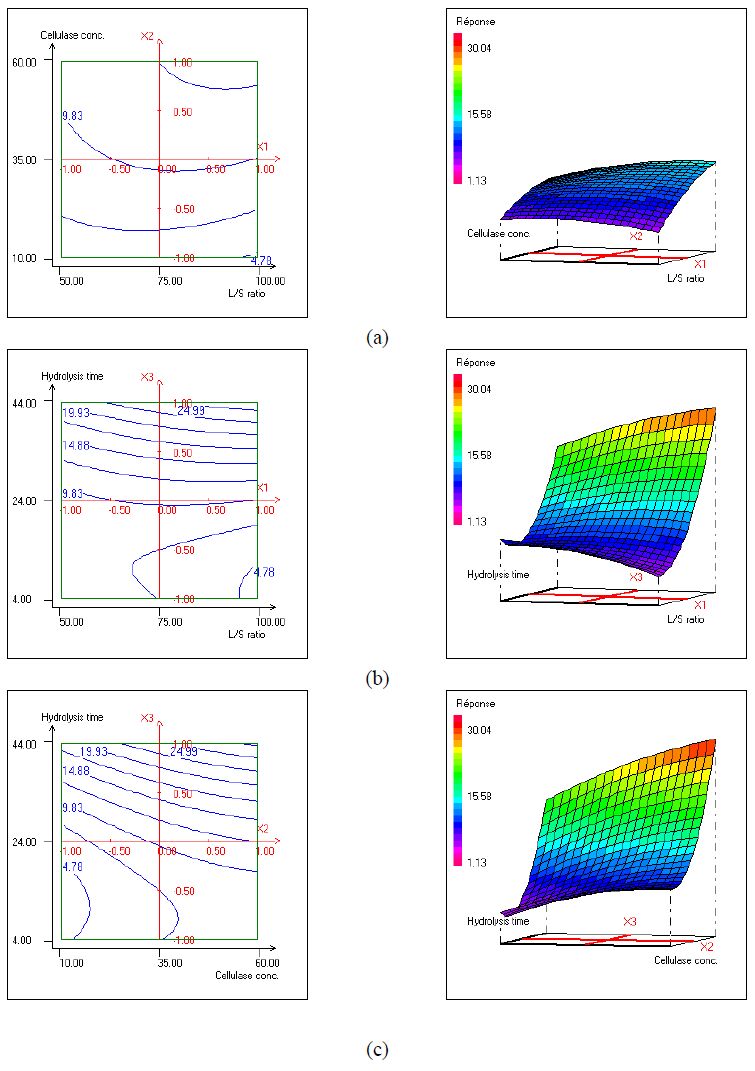
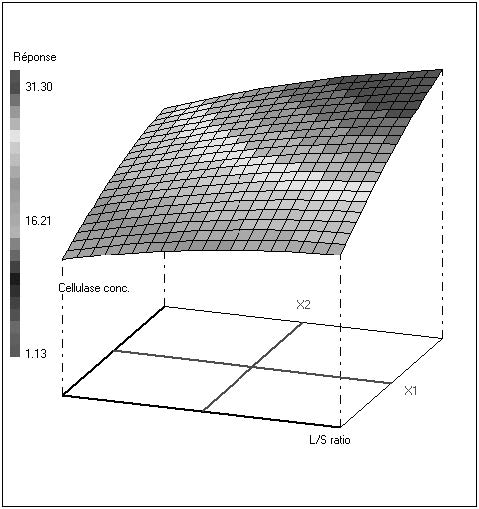
Figures(2) / Tables(5)

Mohamed Neifar, Rim Chatter, Habib Chouchane, Raya Genouiz, Atef Jaouani, Ahmed Slaheddine Masmoudi, Ameur Cherif. Optimization of enzymatic saccharification of Chaetomorpha linum biomass for the production of macroalgae-based third generation bioethanol[J]. AIMS Bioengineering, 2016, 3(3): 400-411. doi: 10.3934/bioeng.2016.3.400







 DownLoad:
DownLoad: