Citation: Khalil Essani, Anton Glieder, Martina Geier. Combinatorial pathway assembly in yeast[J]. AIMS Bioengineering, 2015, 2(4): 423-436. doi: 10.3934/bioeng.2015.4.423

| [1] |
Keasling JD (2012) Synthetic biology and the development of tools for metabolic engineering. Metab Eng 14: 189-195. doi: 10.1016/j.ymben.2012.01.004

|
| [2] |
Stephanopoulos G (2012) Synthetic biology and metabolic engineering. ACS Synth Biol 1: 514-525. doi: 10.1021/sb300094q
|
| [3] |
Yadav V, De Mey M, Lim C, et al. (2012) The future of metabolic engineering and synthetic biology: Towards a systematic practice. Metab Eng 14: 223-241. doi: 10.1016/j.ymben.2011.10.003
|
| [4] | Chao R, Yuan Y, Zhao H (2014) Recent advances in DNA assembly technologies. FEMS Yeast Res 15: 1-9. |
| [5] |
Shetty RP, Endy D, Knight TF (2008) Engineering BioBrick vectors from BioBrick parts. J Biol Eng 2: 5. doi: 10.1186/1754-1611-2-5
|
| [6] |
Engler C, Kandzia R, Marillonnet S (2008) A one pot, one step, precision cloning method with high throughput capability. PLoS One 3: e3647. doi: 10.1371/journal.pone.0003647
|
| [7] |
Gibson D, Young L, Chuang R, et al. (2009) Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat Methods 6: 343-345. doi: 10.1038/nmeth.1318
|
| [8] |
Gibson DG, Benders GA, Axelrod KC, et al. (2008) One-step assembly in yeast of 25 overlapping DNA fragments to form a complete synthetic Mycoplasma genitalium genome. Proc Natl Acad Sci U S A 105: 20404-20409. doi: 10.1073/pnas.0811011106
|
| [9] |
Shao Z, Zhao H, Zhao H (2009) DNA assembler, an in vivo genetic method for rapid construction of biochemical pathways. Nucleic Acids Res 37: e16. doi: 10.1093/nar/gkn724
|
| [10] |
Siddiqui MS, Thodey K, Trenchard I, et al. (2012) Advancing secondary metabolite biosynthesis in yeast with synthetic biology tools. FEMS Yeast Res 12: 144-170. doi: 10.1111/j.1567-1364.2011.00774.x
|
| [11] |
Andersen SL, Sekelsky J (2010) Meiotic versus mitotic recombination: Two different routes for double-strand break repair. Bioessays 32: 1058-1066. doi: 10.1002/bies.201000087
|
| [12] |
Szostak J, Orr-Weaver T, Rothstein R, et al. (1983) The double-strand-break repair model for recombination. Cell 33: 25-35. doi: 10.1016/0092-8674(83)90331-8
|
| [13] |
Aylon Y, Kupiec M (2004) New insights into the mechanism of homologous recombination in yeast. Mutat Res 566: 231-248. doi: 10.1016/j.mrrev.2003.10.001
|
| [14] | Symington LS, Rothstein R, Lisby M (2014) Mechanisms and regulation of mitotic recombination in Saccharomyces cerevisiae. Genetics 198: 795-835. |
| [15] |
Näätsaari L, Mistlberger B, Ruth C, et al. (2012) Deletion of the Pichia pastoris KU70 homologue facilitates platform strain generation for gene expression and synthetic biology. PLoS One 7: e39720. doi: 10.1371/journal.pone.0039720
|
| [16] |
Daley JM, Palmbos PL, Wu D, et al. (2005) Nonhomologous end joining in yeast. Annu Rev Genet 39: 431-451. doi: 10.1146/annurev.genet.39.073003.113340
|
| [17] |
Lorenz M, Muir R, Lim E, et al. (1995) Gene disruption with PCR products in Saccharomyces cerevisiae. Gene 158: 113-117. doi: 10.1016/0378-1119(95)00144-U
|
| [18] |
Manivasakam P, Weber S, McElver J, et al. (1995) Micro-homology mediated PCR targeting in Saccharomyces cerevisiae. Nucleic Acids Res 23: 2799-2800. doi: 10.1093/nar/23.14.2799
|
| [19] | Cregg J, Russell K (1998) Transformation. In: Higgins D, Cregg J (eds) Pichia Protoc. Humana Press Inc, Totowa, NJ, 27-29. |
| [20] | Li P, Anumanthan A, Gao X, et al. (2007) Expression of recombinant proteins in Pichia pastoris. Appl Microbiol Biotechnol 142: 105-124. |
| [21] |
Ma H, Kunes S, Schatz P, et al. (1987) Plasmid construction by homologous recombination in yeast. Gene 58: 201-216. doi: 10.1016/0378-1119(87)90376-3
|
| [22] |
Oldenburg K, Vo K, Michaelis S, et al. (1997) Recombination-mediated PCR-directed plasmid construction in vivo in yeast. Nucleic Acids Res 25: 451-452. doi: 10.1093/nar/25.2.451
|
| [23] |
Shao Z, Luo Y, Zhao H (2012) DNA assembler method for construction of zeaxanthin-producing strains of Saccharomyces cerevisiae. Methods Mol Biol 898: 251-262. doi: 10.1007/978-1-61779-918-1_17
|
| [24] |
Shao Z, Luo Y, Zhao H (2011) Rapid characterization and engineering of natural product biosynthetic pathways via DNA assembler. Mol Biosyst 7: 1056-1059. doi: 10.1039/c0mb00338g
|
| [25] |
Kuijpers NG , Chroumpi S, Vos T, et al. (2013) One-step assembly and targeted integration of multigene constructs assisted by the I-SceI meganuclease in Saccharomyces cerevisiae. FEMS Yeast Res 13: 769-781. doi: 10.1111/1567-1364.12087
|
| [26] |
Wingler LM, Cornish VW (2011) Reiterative recombination for the in vivo assembly of libraries of multigene pathways. Proc Natl Acad Sci U S A 108: 15135-15140. doi: 10.1073/pnas.1100507108
|
| [27] |
Naesby M, Nielsen SV, Nielsen CA, et al. (2009) Yeast artificial chromosomes employed for random assembly of biosynthetic pathways and production of diverse compounds in Saccharomyces cerevisiae. Microb Cell Fact 8: 45. doi: 10.1186/1475-2859-8-45
|
| [28] |
Kim B, Du J, Eriksen DT, et al. (2013) Combinatorial design of a highly efficient xylose-utilizing pathway in Saccharomyces cerevisiae for the production of cellulosic biofuels. Appl Environ Microbiol 79: 931-941. doi: 10.1128/AEM.02736-12
|
| [29] |
Eriksen DT, Hsieh PC, Lynn P, et al. (2013) Directed evolution of a cellobiose utilization pathway in Saccharomyces cerevisiae by simultaneously engineering multiple proteins. Microb Cell Fact 12: 61. doi: 10.1186/1475-2859-12-61
|
| [30] |
Ro DK, Paradise EM, Ouellet M, et al. (2006) Production of the antimalarial drug precursor artemisinic acid in engineered yeast. Nature 440: 940-943. doi: 10.1038/nature04640
|
| [31] |
Westfall PJ, Pitera DJ, Lenihan JR, et al. (2012) Production of amorphadiene in yeast, and its conversion to dihydroartemisinic acid, precursor to the antimalarial agent artemisinin. Proc Natl Acad Sci 109: E111-E118. doi: 10.1073/pnas.1110740109
|
| [32] |
Yamada R, Taniguchi N, Tanaka T, et al. (2010) Cocktail delta-integration: a novel method to construct cellulolytic enzyme expression ratio-optimized yeast strains. Microb Cell Fact 9: 32. doi: 10.1186/1475-2859-9-32
|
| [33] |
Kato H, Matsuda F, Yamada R, et al. (2013) Cocktail δ-integration of xylose assimilation genes for efficient ethanol production from xylose in Saccharomyces cerevisiae. J Biosci Bioeng 116: 333-336. doi: 10.1016/j.jbiosc.2013.03.020
|
| [34] | Yuan J, Ching CB (2014) Combinatorial assembly of large biochemical pathways into yeast chromosomes for improved production of value-added compounds. ACS Synth Biol 4: 23-31. |
| [35] |
Sakai A, Shimizu Y, Hishinuma F (1990) Integration of heterologous genes into the chromosome of Saccharomyces cerevisiae using a delta sequence of yeast retrotransposon Ty. Appl Microbiol Biotechnol 33: 302-306. doi: 10.1007/BF00164526
|
| [36] |
Du J, Yuan Y, Si T, et al. (2012) Customized optimization of metabolic pathways by combinatorial transcriptional engineering. Nucleic Acids Res 40:e142. doi: 10.1093/nar/gks549
|
| [37] |
Mitchell LA, Chuang J, Agmon N, et al. (2015) Versatile genetic assembly system (VEGAS) to assemble pathways for expression in S. cerevisiae. Nucleic Acids Res 43: 6620-6630. doi: 10.1093/nar/gkv466
|
| [38] |
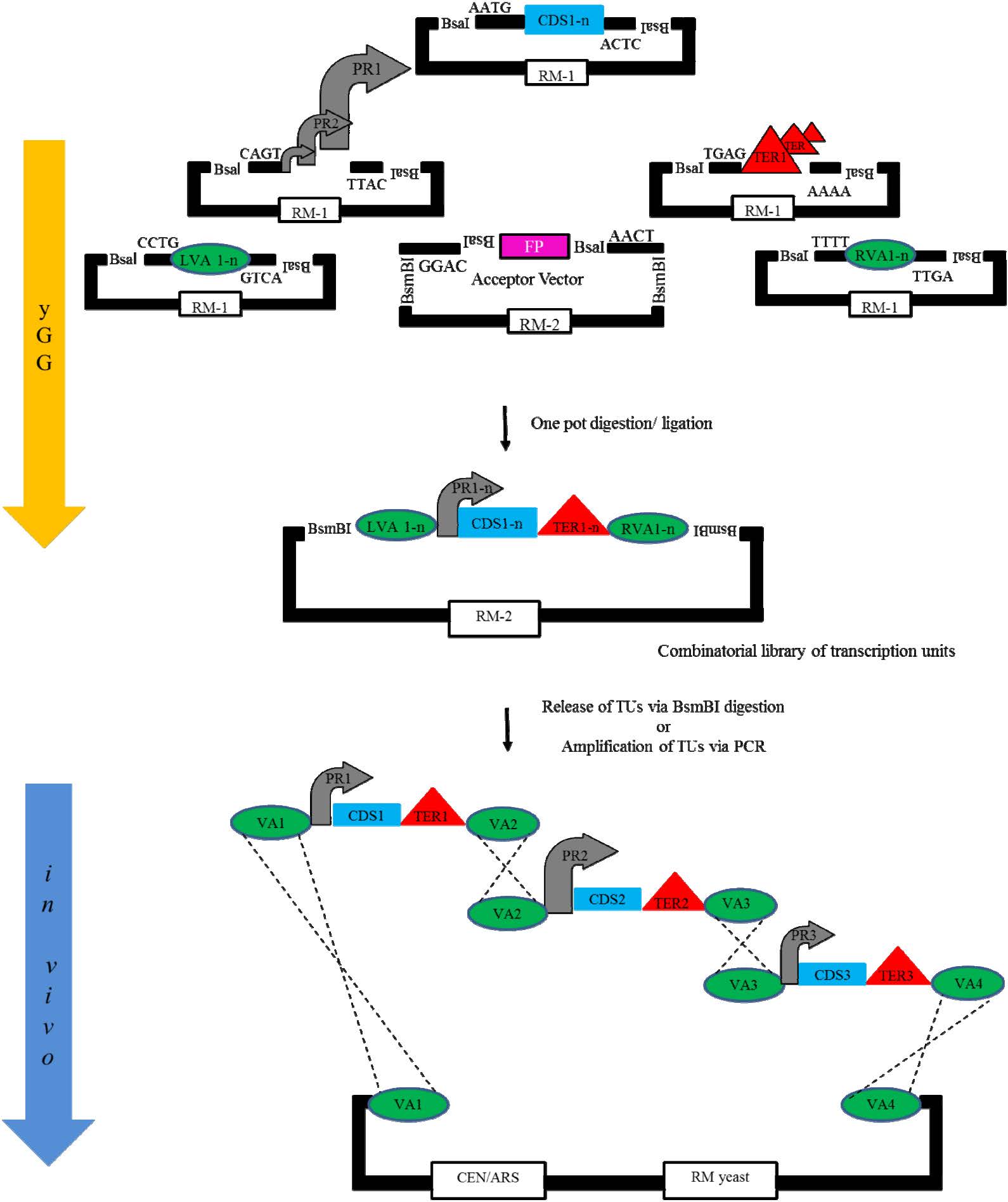
Agmon N, Mitchell LA., Cai Y, et al. (2015) Yeast Golden Gate (yGG) for efficient assembly of S. cerevisiae transcription units. ACS Synth Biol 4: 853-859. doi: 10.1021/sb500372z
|
| [39] |
Mézard C, Pompon D, Nicolas A (1992) Recombination between similar but not identical DNA sequences during yeast transformation occurs within short stretches of identity. Cell 70:659-670. doi: 10.1016/0092-8674(92)90434-E
|
| [40] | Abécassis V, Pompon D, Truan G (2000) High efficiency family shuffling based on multi-step PCR and in vivo DNA recombination in yeast: statistical and functional analysis of a combinatorial library between human cytochrome P450 1A1 and 1A2. Nucleic Acids Res 28: E88. |
| [41] |
Swers J, Kellogg B, Wittrup K (2004) Shuffled antibody libraries created by in vivo homologous recombination and yeast surface display. Nucleic Acids Res 32:e36. doi: 10.1093/nar/gnh030
|
| [42] |
Luque A, Sebai SC, Santiago-Schübel B, et al. (2014) In vivo evolution of metabolic pathways by homeologous recombination in mitotic cells. Metab Eng 23: 123-135. doi: 10.1016/j.ymben.2014.02.010
|
| [43] |
Beekwilder J, van Rossum HM, Koopman F, et al. (2014) Polycistronic expression of a β-carotene biosynthetic pathway in Saccharomyces cerevisiae coupled to β-ionone production. J Biotechnol 192: 383-392. doi: 10.1016/j.jbiotec.2013.12.016
|
| [44] | Geier M, Fauland PC, Vogl T, et al. (2014) Compact multi enzyme pathways in P. pastoris. Chem Commun 51: 1643-1646. |
| [45] |
De Felipe P, Luke GA, Hughes LE, et al. (2006) E unum pluribus: multiple proteins from a self-processing polyprotein. Trends Biotechnol 24: 68-75. doi: 10.1016/j.tibtech.2005.12.006
|
| [46] |
Pfleger BF, Pitera DJ, Smolke CD, et al. (2006) Combinatorial engineering of intergenic regions in operons tunes expression of multiple genes. Nat Biotechnol 24: 1027-1032. doi: 10.1038/nbt1226
|
Figures(4) / Tables(1)

Khalil Essani, Anton Glieder, Martina Geier. Combinatorial pathway assembly in yeast[J]. AIMS Bioengineering, 2015, 2(4): 423-436. doi: 10.3934/bioeng.2015.4.423







 DownLoad:
DownLoad: