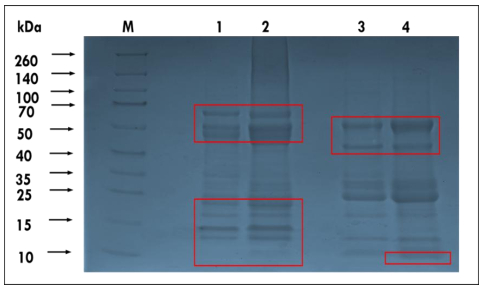
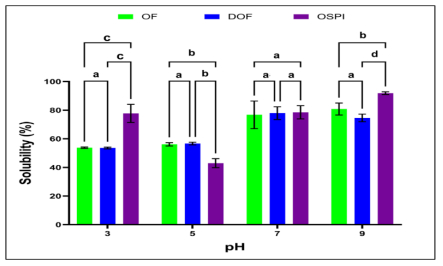
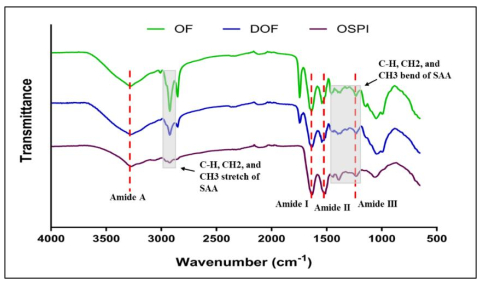
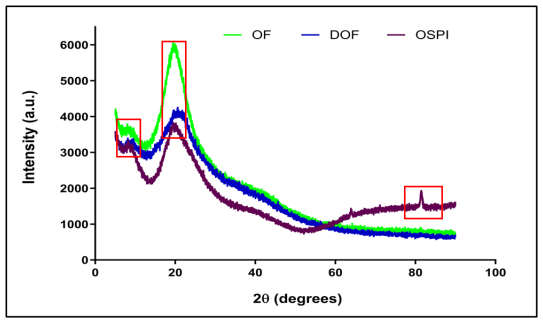
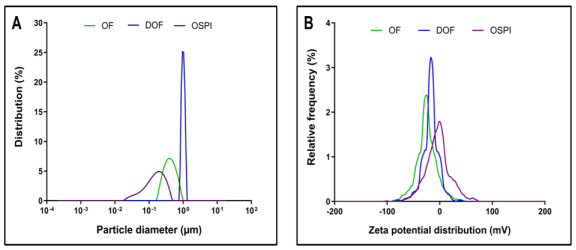
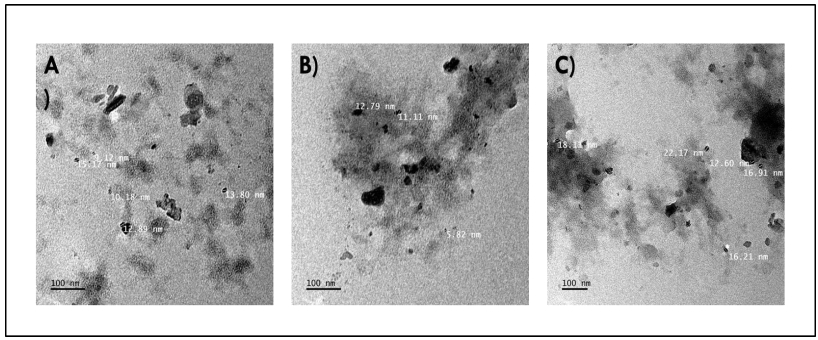
Okra seed protein isolate (OSPI) has garnered increasing interest due to its bioactive potential and nutritional value. This research focused on analyzing the structural characteristics and molecular composition of OSPI to evaluate its potential therapeutic applications. The amino acid profile revealed a diverse range of amino acids, with leucine (6.56 g/100 g), glutamic acid (14.46 g/100 g), and arginine (9.42 g/100 g) identified as the most abundant. Hydrophobic amino acids (31.39 g/100 g) along with branched-chain amino acids (14.24 g/100 g) further underscore their bioactivity, particularly in immune modulation and cancer therapy. Protein solubility analysis indicated optimal solubility (91.86%) at pH 9, a key factor influencing bioavailability. The molecular weight profile identified polypeptide bands corresponding to 2S albumins, known for their antimicrobial and immunomodulatory properties. FTIR and XRD analyses highlighted the presence of α-helical and β-sheet structures, contributing to OSPI's functional stability. Additionally, OSPI exhibited an amorphous nature, which is beneficial for pharmaceutical applications due to enhanced bioavailability. Particle size analysis demonstrated a monomodal distribution, with OSPI displaying the smallest size (0.144 ± 0.1 µm), indicative of improved drug delivery potential. Collectively, these findings emphasize OSPI's therapeutic potential, particularly in disease prevention and treatment, supporting its potential application in pharmaceutical and nutraceutical industries.
Citation: Letisha Maistry, Abe Shegro Gerrano, John Mellem. Chemical and structural characterization of okra (Abelmoschus esculentus) seed protein isolate as a potential therapeutic agent[J]. AIMS Agriculture and Food, 2025, 10(2): 353-370. doi: 10.3934/agrfood.2025018
Okra seed protein isolate (OSPI) has garnered increasing interest due to its bioactive potential and nutritional value. This research focused on analyzing the structural characteristics and molecular composition of OSPI to evaluate its potential therapeutic applications. The amino acid profile revealed a diverse range of amino acids, with leucine (6.56 g/100 g), glutamic acid (14.46 g/100 g), and arginine (9.42 g/100 g) identified as the most abundant. Hydrophobic amino acids (31.39 g/100 g) along with branched-chain amino acids (14.24 g/100 g) further underscore their bioactivity, particularly in immune modulation and cancer therapy. Protein solubility analysis indicated optimal solubility (91.86%) at pH 9, a key factor influencing bioavailability. The molecular weight profile identified polypeptide bands corresponding to 2S albumins, known for their antimicrobial and immunomodulatory properties. FTIR and XRD analyses highlighted the presence of α-helical and β-sheet structures, contributing to OSPI's functional stability. Additionally, OSPI exhibited an amorphous nature, which is beneficial for pharmaceutical applications due to enhanced bioavailability. Particle size analysis demonstrated a monomodal distribution, with OSPI displaying the smallest size (0.144 ± 0.1 µm), indicative of improved drug delivery potential. Collectively, these findings emphasize OSPI's therapeutic potential, particularly in disease prevention and treatment, supporting its potential application in pharmaceutical and nutraceutical industries.

| [1] |
Kumar M, Tomar M, Punia S, et al. (2022) Plant-based proteins and their multifaceted industrial applications. LWT 154: 112620. https://doi.org/10.1016/j.lwt.2021.112620 doi: 10.1016/j.lwt.2021.112620

|
| [2] |
Li C, Yang J, Yao L, et al. (2020) Characterisation, physicochemical and functional properties of protein isolates from Amygdalus pedunculata pall seeds. Food Chem 311: 125888. https://doi.org/10.1016/j.foodchem.2019.125888 doi: 10.1016/j.foodchem.2019.125888
|
| [3] |
Sim SYJ, Srv A, Chiang JH, et al. (2021) Plant proteins for future foods: A roadmap. Foods 10: 1967. https://doi.org/10.3390/foods10081967 doi: 10.3390/foods10081967
|
| [4] |
Tan M, Nawaz MA, Buckow R (2023) Functional and food application of plant proteins–a review. Food Rev Int 39: 2428–2456. https://doi.org/10.1080/87559129.2021.1955918 doi: 10.1080/87559129.2021.1955918
|
| [5] |
Bhutia SK, Panda PK, Sinha N, et al. (2019) Plant lectins in cancer therapeutics: Targeting apoptosis and autophagy-dependent cell death. Pharmacol Res 144: 8–18. https://doi.org/10.1016/j.phrs.2019.04.001 doi: 10.1016/j.phrs.2019.04.001
|
| [6] |
Tomar M, Bhardwaj R, Kumar M, et al. (2021) Nutritional composition patterns and application of multivariate analysis to evaluate indigenous Pearl millet ((Pennisetum glaucum (L.) R. Br.) germplasm. J Food Compos Anal 103: 104086. https://doi.org/10.1016/j.jfca.2021.104086 doi: 10.1016/j.jfca.2021.104086
|
| [7] | Ansori ANM (2021) A mini-review of the medicinal properties of Okra (Abelmoschus esculentus l.) and potential benefit against SARS-CoV-2. Indian J Forensic Med Toxicol 15: 852–856. |
| [8] |
Shahnaz T, Fawole AO, Adeyanju AA, et al. (2024) Food proteins as functional ingredients in the management of chronic diseases: A concise review. Nutrients 16: 2323. https://doi.org/10.3390/nu16142323 doi: 10.3390/nu16142323
|
| [9] |
Peighambardoust SH, Karami Z, Pateiro M, et al. (2021) A review on health-promoting, biological, and functional aspects of bioactive peptides in food applications. Biomolecules 11: 631. https://doi.org/10.3390/biom11050631 doi: 10.3390/biom11050631
|
| [10] | Bonciu E (2020) Aspects of the involvement of biotechnology in functional food and nutraceuticals. Available from: https://agronomyjournal.usamv.ro/pdf/2020/issue_2/Art43.pdf. |
| [11] |
Ofori J, Tortoe C, Agbenorhevi JK (2020) Physicochemical and functional properties of dried okra (Abelmoschus esculentus L.) seed flour. Food Sci Nutr 8: 4291–4296. https://doi.org/10.1002/fsn3.1725 doi: 10.1002/fsn3.1725
|
| [12] |
Elkhalifa AEO, Alshammari E, Adnan M, et al. (2021) Okra (Abelmoschus esculentus) as a potential dietary medicine with nutraceutical importance for sustainable health applications. Molecules 26: 696. https://doi.org/10.3390/molecules26030696 doi: 10.3390/molecules26030696
|
| [13] |
Esmaeilzadeh D, Razavi BM, Hosseinzadeh H (2020) Effect of Abelmoschus esculentus (okra) on metabolic syndrome: A review. Phytother Res 34: 2192–2202. https://doi.org/10.1002/ptr.6679 doi: 10.1002/ptr.6679
|
| [14] |
Liu Y, Qi J, Luo J, et al. (2021) Okra in food field: nutritional value, health benefits and effects of processing methods on quality. Food Rev Int 37: 67–90. https://doi.org/10.1080/87559129.2019.1695833 doi: 10.1080/87559129.2019.1695833
|
| [15] | Gao H, Zhang W, Wang B, et al. (2018) Purification, characterization and anti-fatigue activity of polysaccharide fractions from okra (Abelmoschus esculentus (L.) Moench). Food & Function 9: 1088–1101. https://pubs.rsc.org/en/content/articlehtml/2018/fo/c7fo01821e |
| [16] |
Xia F, Zhong Y, Li M, et al. (2015) Antioxidant and anti-fatigue constituents of okra. Nutrients 7: 8846-8858.. https://doi.org/10.3390/nu7105435 doi: 10.3390/nu7105435
|
| [17] |
Yuan Q, Lin S, Fu Y, et al. (2019) Effects of extraction methods on the physicochemical characteristics and biological activities of polysaccharides from okra (Abelmoschus esculentus). Int J Bio Macromol127: 178–186. https://doi.org/10.1016/j.ijbiomac.2019.01.042 doi: 10.1016/j.ijbiomac.2019.01.042
|
| [18] |
Kahlon T, Chapman M, Smith G (2007) In vitro binding of bile acids by okra, beets, asparagus, eggplant, turnips, green beans, carrots, and cauliflower. Food Chem 103: 676–680. https://doi.org/10.1016/j.foodchem.2006.07.056 doi: 10.1016/j.foodchem.2006.07.056
|
| [19] | Petropoulos S, Fernandes A, Barros L, et al. (2017) The chemical composition, nutritional value and antimicrobial properties of Abelmoschus esculentus seeds. Food Funct 8: 4733–4743. https://pubs.rsc.org/en/content/articlehtml/2017/fo/c7fo01446e |
| [20] |
Mairuae N, Cheepsunthorn P, Cheepsunthorn CL, et al. (2017) Okra (Abelmoschus esculentus Linn) inhibits lipopolysaccharide-induced inflammatory mediators in BV2 microglial cells. Trop J Pharm Res 16: 1285–1292. https://doi.org/10.4314/tjpr.v16i6.11 doi: 10.4314/tjpr.v16i6.11
|
| [21] |
Zheng W, Zhao T, Feng W, et al. (2014) Purification, characterization and immunomodulating activity of a polysaccharide from flowers of Abelmoschus esculentus. Carbohyd Polym 106: 335–342. https://doi.org/10.1016/j.carbpol.2014.02.079 doi: 10.1016/j.carbpol.2014.02.079
|
| [22] |
Monte LG, Santi-Gadelha T, Reis LB, et al. (2014) Lectin of Abelmoschus esculentus (okra) promotes selective antitumor effects in human breast cancer cells. Biotechnol Lett 36: 461–469. https://doi.org/10.1007/s10529-013-1382-4 doi: 10.1007/s10529-013-1382-4
|
| [23] |
Nnamezie AA, Famuwagun AA, Gbadamosi SO (2021) Characterization of okra seed flours, protein concentrate, protein isolate and enzymatic hydrolysates. Food Prod, Process Nutr 3: 14. https://doi.org/10.1186/s43014-021-00059-9 doi: 10.1186/s43014-021-00059-9
|
| [24] |
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72: 248–254. https://doi.org/10.1016/0003-2697(76)90527-3 doi: 10.1016/0003-2697(76)90527-3
|
| [25] |
Chen Z, Wang J, Liu W, et al. (2017) Physicochemical characterization, antioxidant and anticancer activities of proteins from four legume species. J Food Sci Technol 54: 964–972. https://doi.org/10.1007/s13197-016-2390-x doi: 10.1007/s13197-016-2390-x
|
| [26] | Wheat TE, Grumbach ES, Mazzeo JR (2008) UPLC amino acid analysis solution. Available from: https://www.waters.com/webassets/cms/library/docs/720001683en.pdf. |
| [27] |
Madurapperumage A, Johnson N, Thavarajah P, et al. (2022) Fourier‐transform infrared spectroscopy (FTIR) as a high‐throughput phenotyping tool for quantifying protein quality in pulse crops. Plant Phenome J 5: e20047. https://doi.org/10.1002/ppj2.20047 doi: 10.1002/ppj2.20047
|
| [28] |
Sahni P, Sharma S, Surasani VKR (2020) Influence of processing and pH on amino acid profile, morphology, electrophoretic pattern, bioactive potential and functional characteristics of alfalfa protein isolates. Food Chem 333: 127503. https://doi.org/10.1016/j.foodchem.2020.127503 doi: 10.1016/j.foodchem.2020.127503
|
| [29] |
Alabi OO, Annor GA, Amonsou EO (2023) Effect of cold plasma-activated water on the physicochemical and functional properties of Bambara groundnut globulin. Food Struct 36: 100321. https://doi.org/10.1016/j.foostr.2023.100321 doi: 10.1016/j.foostr.2023.100321
|
| [30] |
Du C, Wang Z, Zheng Z (2023) The preparation of plant-based milk substitutes with antioxidant properties using soybean protein isolate and curcumin composite nanoparticles. LWT 182: 114780. https://doi.org/10.1016/j.lwt.2023.114780 doi: 10.1016/j.lwt.2023.114780
|
| [31] |
Ling Z-N, Jiang Y-F, Ru J-N, et al. (2023) Amino acid metabolism in health and disease. Signal Transduct Tar Ther 8: 345. https://doi.org/10.1038/s41392-023-01569-3 doi: 10.1038/s41392-023-01569-3
|
| [32] |
Ahmed N, Ali A, Riaz S, et al. (2021) Vegetable proteins: Nutritional value, sustainability, and future perspectives. Veg Crops-Health Benefits Cultiv: 1–14. https://doi.org/10.5772/intechopen.101126 doi: 10.5772/intechopen.101126
|
| [33] |
Akbay B, Omarova Z, Trofimov A, et al. (2024) Double-Edge Effects of Leucine on Cancer Cells. Biomolecules 14: 1401. https://doi.org/10.3390/biom14111401 doi: 10.3390/biom14111401
|
| [34] |
Ijarotimi OS, Akinola-Ige AO, Oluwajuyitan TD (2023) Okra seeds proteins: Amino acid profile, free radical scavenging activities and inhibition of diabetes and hypertensive converting enzymes indices. Meas: Food 11: 100101. https://doi.org/10.1016/j.meafoo.2023.100101 doi: 10.1016/j.meafoo.2023.100101
|
| [35] |
Chen J, Cui L, Lu S, et al. (2024) Amino acid metabolism in tumor biology and therapy. Cell Death Dis 15: 42. https://doi.org/10.1038/s41419-024-06435-w doi: 10.1038/s41419-024-06435-w
|
| [36] |
Dai Y, Cai X, Shi W, et al. (2017) Pro-apoptotic cationic host defense peptides rich in lysine or arginine to reverse drug resistance by disrupting tumor cell membrane. Amino Acids 49: 1601–1610. https://doi.org/10.1007/s00726-017-2453-y doi: 10.1007/s00726-017-2453-y
|
| [37] |
Yamaguchi Y, Yamamoto K, Sato Y, et al. (2016) Combination of aspartic acid and glutamic acid inhibits tumor cell proliferation. Biomed Res 37: 153–159. https://doi.org/10.2220/biomedres.37.153 doi: 10.2220/biomedres.37.153
|
| [38] |
Chalamaiah M, Yu W, Wu J (2018) Immunomodulatory and anticancer protein hydrolysates (peptides) from food proteins: A review. Food Chem 245: 205–222. https://doi.org/10.1016/j.foodchem.2017.10.087 doi: 10.1016/j.foodchem.2017.10.087
|
| [39] |
Wang Z, Zhang X, Wang L, et al. (2024) High Fischer ratio oligopeptides in food: sources, functions and application prospects. J Future Foods 4: 128–134. https://doi.org/10.1016/j.jfutfo.2023.06.003 doi: 10.1016/j.jfutfo.2023.06.003
|
| [40] | FAO/WHO/UNU (1985) Energy and protein requirements, report of the joint FAO/WHO/UNU expert consultation. Technical Report series No. 724. Available from: https://scholar.google.com/scholar?hl=en&as_sdt=0%2C5&q=FAO%2FWHO%2FUNU+%281985%29+Energy+and+protein+requirements%2C+report+of+the+joint+FAO%2FWHO%2FUNU+expert+consultation.Technical+Report+series+No.+724.+Geneva%2C+Switzerland%3A+FAO%2C+WHO+and+the+United+Nations+University. & btnG = . |
| [41] |
Patil PJ, Usman M, Zhang C, et al. (2022) An updated review on food-derived bioactive peptides: Focus on the regulatory requirements, safety, and bioavailability. Compr Rev Food Sci Food Saf 21: 1732–1776. https://doi.org/10.1111/1541-4337.12911 doi: 10.1111/1541-4337.12911
|
| [42] |
Naiker TS, Baijnath H, Amonsou EO, et al. (2020) Effect of steaming and dehydration on the nutritional quality and functional properties of protein isolates produced from Lablab purpureus (L.) Sweet (hyacinth bean). J Food Process Preserv 44: e14334. https://doi.org/10.1111/jfpp.14334 doi: 10.1111/jfpp.14334
|
| [43] |
Fang C, Kanemaru K, Carvalho WSP, et al. (2024) Self-assembled poloxamer-legumin/vicilin nanoparticles for the nanoencapsulation and controlled release of folic acid. Int J Bio Macromol 268: 131646. https://doi.org/10.1016/j.ijbiomac.2024.131646 doi: 10.1016/j.ijbiomac.2024.131646
|
| [44] | Antonia V, Vera A, Manuel V, et al. (2019) Conformational and physicochemical properties of quinoa proteins affected by different conditions of high-intensity ultrasound treatments. Ultrason Sonochem 51: 186–196. https://pubmed.ncbi.nlm.nih.gov/30377080 |
| [45] |
Souza PFN (2020) The forgotten 2S albumin proteins: Importance, structure, and biotechnological application in agriculture and human health. Int J Bio Macromol 164: 4638–4649. https://doi.org/10.1016/j.ijbiomac.2020.09.049 doi: 10.1016/j.ijbiomac.2020.09.049
|
| [46] |
Laxminarayan R, Duse A, Wattal C, et al. (2013) Antibiotic resistance—the need for global solutions. Lancet Infect Dis 13: 1057–1098. https://doi.org/10.1016/S1473-3099(13)70318-9 doi: 10.1016/S1473-3099(13)70318-9
|
| [47] |
Maria-Neto S, Honorato RV, Costa FT, et al. (2011) Bactericidal Activity Identified in 2S Albumin from Sesame Seeds and In silico Studies of Structure–Function Relations. Protein J 30: 340–350. https://doi.org/10.1007/s10930-011-9337-x doi: 10.1007/s10930-011-9337-x
|
| [48] |
González-Montoya M, Hernández-Ledesma B, Silván JM, et al. (2018) Peptides derived from in vitro gastrointestinal digestion of germinated soybean proteins inhibit human colon cancer cells proliferation and inflammation. Food Chem 242: 75–82. https://doi.org/10.1016/j.foodchem.2017.09.035 doi: 10.1016/j.foodchem.2017.09.035
|
| [49] | Hu S, Liu C, Liu X (2023) The beneficial effects of soybean proteins and peptides on chronic diseases. Nutrients 15: 1811. https://www.mdpi.com/2072-6643/15/8/1811 |
| [50] |
Marcela G-M, Eva R-G, Del Carmen R-RM, et al. (2016) Evaluation of the antioxidant and antiproliferative effects of three peptide fractions of germinated soybeans on breast and cervical cancer cell lines. Plant Foods Human Nutr 71: 368–374. https://doi.org/10.1007/s11130-016-0568-z doi: 10.1007/s11130-016-0568-z
|
| [51] |
Chen J, Liu G, Pantalone V, et al. (2020) Physicochemical properties of proteins extracted from four new Tennessee soybean lines. J Agric Food Res 2: 100022. https://doi.org/10.1016/j.jafr.2020.100022 doi: 10.1016/j.jafr.2020.100022
|
| [52] | Ivanova P, Chalova V, Koleva L, et al. (2013) Amino acid composition and solubility of proteins isolated from sunflower meal produced in Bulgaria. Int Food Res J 20: 2995–3000. |
| [53] |
Yagasaki K, Takagi T, Sakai M, et al. (1997) Biochemical characterization of soybean protein consisting of different subunits of glycinin. J Agric Food Chem 45: 656–660. https://doi.org/10.1021/jf9604394 doi: 10.1021/jf9604394
|
| [54] |
Mir NA, Riar CS, Singh S (2019) Effect of pH and holding time on the characteristics of protein isolates from Chenopodium seeds and study of their amino acid profile and scoring. Food Chem 272: 165–173. https://doi.org/10.1016/j.foodchem.2018.08.048 doi: 10.1016/j.foodchem.2018.08.048
|
| [55] |
Zhang L, Xiao Q, Wang Y, et al. (2017) Denatured protein-coated docetaxel nanoparticles: Alterable drug state and cytosolic delivery. Int J Pharm 523: 1–14. https://doi.org/10.1016/j.ijpharm.2017.03.026 doi: 10.1016/j.ijpharm.2017.03.026
|
| [56] |
Sadat A, Joye IJ (2020) Peak fitting applied to fourier transform infrared and raman spectroscopic analysis of proteins. Appl Sci 10: 5918. https://doi.org/10.3390/app10175918 doi: 10.3390/app10175918
|
| [57] |
Krimm S, Bandekar J (1986) Vibrational spectroscopy and conformation of peptides, polypeptides, and proteins. Adv Protein Chem 38: 181–364. https://doi.org/10.1016/S0065-3233(08)60528-8 doi: 10.1016/S0065-3233(08)60528-8
|
| [58] | Aboul-eneın Y, Bunacıu A, Fleschın S (2014) Evaluation of the protein secondary structures using Fourier transform infrared spectroscopy. Gazi Univ J Sci 27: 637–644. https://dergipark.org.tr/en/pub/gujs/issue/7430/97685 |
| [59] |
Kong J, Yu S (2007) Fourier Transform Infrared Spectroscopic Analysis of Protein Secondary Structures. Acta Bioch Bioph Sin 39: 549–559. https://doi.org/10.1111/j.1745-7270.2007.00320.x doi: 10.1111/j.1745-7270.2007.00320.x
|
| [60] |
Malik MA, Sharma HK, Saini CS (2016) Effect of removal of phenolic compounds on structural and thermal properties of sunflower protein isolate. J Food Sci Technol 53: 3455–3464. https://doi.org/10.1007/s13197-016-2320-y doi: 10.1007/s13197-016-2320-y
|
| [61] |
Aitipamula S, Vangala VR (2017) X-ray crystallography and its role in understanding the physicochemical properties of pharmaceutical cocrystals. J Indian Inst Sci 97: 227–243. https://doi.org/10.1007/s41745-017-0026-4 doi: 10.1007/s41745-017-0026-4
|
| [62] | Jhan F, Gani A, Noor N, et al. (2021) Nanoreduction of millet proteins: Effect on structural and functional properties. ACS Food Sci Technol 1: 1418–1427. https://pubs.acs.org/doi/abs/10.1021/acsfoodscitech.1c00132 |
| [63] |
Khan WA, Butt MS, Pasha I, et al. (2020) Microencapsulation of vitamin D in protein matrices: In vitro release and storage stability. J Food Meas Charact 14: 1172–1182. https://doi.org/10.1007/s11694-019-00366-3 doi: 10.1007/s11694-019-00366-3
|
| [64] |
Hoseini B, Jaafari MR, Golabpour A, et al. (2023) Application of ensemble machine learning approach to assess the factors affecting size and polydispersity index of liposomal nanoparticles. Sci Rep 13: 18012. https://doi.org/10.1038/s41598-023-43689-4 doi: 10.1038/s41598-023-43689-4
|
| [65] |
Danaei M, Dehghankhold M, Ataei S, et al. (2018) Impact of particle size and polydispersity index on the clinical applications of lipidic nanocarrier systems. Pharmaceutics 10: 57. https://doi.org/10.3390/pharmaceutics10020057 doi: 10.3390/pharmaceutics10020057
|
| [66] |
Németh Z, Csóka I, Semnani Jazani R, et al. (2022) Quality by design-driven zeta potential optimisation study of liposomes with charge imparting membrane additives. Pharmaceutics 14: 1798. https://doi.org/10.3390/pharmaceutics14091798 doi: 10.3390/pharmaceutics14091798
|
| [67] | Thakur P, Sonawane SS, Sonawane SH, et al. (2020) Nanofluids-based delivery system, encapsulation of nanoparticles for stability to make stable nanofluids, In: Encapsulation of Active Molecules and their Delivery System, Elsevier, 141–152. https://doi.org/10.1016/B978-0-12-819363-1.00009-0 |
| [68] |
You JY, Kang SJ, Rhee WJ (2021) Isolation of cabbage exosome-like nanovesicles and investigation of their biological activities in human cells. Bioact Mater 6: 4321–4332. https://doi.org/10.1016/j.bioactmat.2021.04.023 doi: 10.1016/j.bioactmat.2021.04.023
|
| [69] | Sung JJ, Pardeshi NN, Mulder AM, et al. (2015) Transmission electron microscopy as an orthogonal method to characterize protein aggregates. J Pharm Sci 104: 750–759. |
| [70] |
Priscila Laiz Z, Soraia P, Sofia D, et al. (2023) Zein nanoparticles as oral carrier for mometasone furoate delivery. Drug Deliv Transl Res 13: 2948–2959. https://doi.org/10.1007/s13346-023-01367-y doi: 10.1007/s13346-023-01367-y
|
Figures(6) / Tables(1)

Letisha Maistry, Abe Shegro Gerrano, John Mellem. Chemical and structural characterization of okra (Abelmoschus esculentus) seed protein isolate as a potential therapeutic agent[J]. AIMS Agriculture and Food, 2025, 10(2): 353-370. doi: 10.3934/agrfood.2025018







 DownLoad:
DownLoad: