Citation: Georgia Theocharopoulou. The ubiquitous role of mitochondria in Parkinson and other neurodegenerative diseases[J]. AIMS Neuroscience, 2020, 7(1): 43-65. doi: 10.3934/Neuroscience.2020004

| [1] | Gonatas N, Shy G (1965) Childhood myopathies with abnormal mitochondria. Proceedings of the Vth International Congress of Neuropathology 100: 606-612. |
| [2] |
Baron M, Kudin AP, Kunz WS (2007) Mitochondrial dysfunction in neurodegenerative disorders. Biochem Soc Trans 35: 1228-1231. doi: 10.1042/BST0351228

|
| [3] |
DiMauro S (2011) A history of mitochondrial diseases. J Inherit Metab Dis 34: 261-276. doi: 10.1007/s10545-010-9082-x
|
| [4] |
Mattson MP, Gleichmann M, Cheng A (2008) Mitochondria in neuroplasticity and neurological disorders. Neuron 60: 748-766. doi: 10.1016/j.neuron.2008.10.010
|
| [5] |
Federico A, Cardaioli E, Da Pozzo P, et al. (2012) Mitochondria, oxidative stress and neurodegeneration. J Neurol Sci 322: 254-262. doi: 10.1016/j.jns.2012.05.030
|
| [6] |
De Vos KJ, Grierson AJ, Ackerley S, et al. (2008) Role of axonal transport in neurodegenerative diseases. Annu Rev Neurosci 31: 151-173. doi: 10.1146/annurev.neuro.31.061307.090711
|
| [7] |
Chen H, Chan DC (2009) Mitochondrial dynamics–fusion, fission, movement, and mitophagy–in neurodegenerative diseases. Hum Mol Genet 18: R169-R176. doi: 10.1093/hmg/ddp326
|
| [8] |
Wang X, Su B, Zheng L, et al. (2009) The role of abnormal mitochondrial dynamics in the pathogenesis of Alzheimer's disease. J Neurochem 109: 153-159. doi: 10.1111/j.1471-4159.2009.05867.x
|
| [9] |
Büeler H (2009) Impaired mitochondrial dynamics and function in the pathogenesis of Parkinson's disease. Exp Neurol 218: 235-246. doi: 10.1016/j.expneurol.2009.03.006
|
| [10] |
Park JS, Davis RL, Sue CM (2018) Mitochondrial dysfunction in Parkinson's disease: new mechanistic insights and therapeutic perspectives. Curr Neurol Neurosci Rep 18: 21. doi: 10.1007/s11910-018-0829-3
|
| [11] | Alexiou A, Vlamos P, Volikas K (2010) A Theoretical Artificial Approach on Reducing Mitochondrial Abnormalities in Alzheimer Disease. Proceedings of the 10th IEEE International Conference on Information Technology and Applications in Biomedicin IEEE, 1-4. |
| [12] |
Benard G, Karbowski M (2009) Mitochondrial fusion and division: regulation and role in cell viability. Semin Cell Dev Biol 20: 365-374. doi: 10.1016/j.semcdb.2008.12.012
|
| [13] |
Zorov DB, Vorobjev IA, Popkov VA, et al. (2019) Lessons from the Discovery of Mitochondrial Fragmentation (Fission): A Review and Update. Cells 8: 175. doi: 10.3390/cells8020175
|
| [14] |
Sheng ZH (2017) The interplay of axonal energy homeostasis and mitochondrial trafficking and anchoring. Trends Cell Biol 27: 403-416. doi: 10.1016/j.tcb.2017.01.005
|
| [15] |
Lin MY, Sheng ZH (2015) Regulation of mitochondrial transport in neurons. Expe Cell Res 334: 35-44. doi: 10.1016/j.yexcr.2015.01.004
|
| [16] |
Mishra P, Chan DC (2016) Metabolic regulation of mitochondrial dynamics. J Cell Biol 212: 379-387. doi: 10.1083/jcb.201511036
|
| [17] |
Dickey AS, Strack S (2011) PKA/AKAP1 and PP2A/Bβ2 regulate neuronal morphogenesis via Drp1 phosphorylation and mitochondrial bioenergetics. J Neurosci 31: 15716-15726. doi: 10.1523/JNEUROSCI.3159-11.2011
|
| [18] |
Kuznetsov AV, Hermann M, Saks V, et al. (2009) The cell-type specificity of mitochondrial dynamics. Int J Biochem Cell B 41: 1928-1939. doi: 10.1016/j.biocel.2009.03.007
|
| [19] |
Palmer CS, Osellame LD, Stojanovski D, et al. (2011) The regulation of mitochondrial morphology: intricate mechanisms and dynamic machinery. Cell Signal 23: 1534-1545. doi: 10.1016/j.cellsig.2011.05.021
|
| [20] |
Chen H, McCaffery JM, Chan DC (2007) Mitochondrial Fusion Protects against Neurodegeneration in the Cerebellum. Cell 130: 548-562. doi: 10.1016/j.cell.2007.06.026
|
| [21] |
Twig G, Elorza A, Molina AJ, et al. (2008) Fission and selective fusion govern mitochondrial segregation and elimination by autophagy. EMBO J 27: 433-446. doi: 10.1038/sj.emboj.7601963
|
| [22] |
Meeusen S, DeVay R, Block J, et al. (2006) Mitochondrial inner-membrane fusion and crista maintenance requires the dynamin-related GTPase Mgm1. Cell 127: 383-395. doi: 10.1016/j.cell.2006.09.021
|
| [23] |
Liesa M, Palacín M, Zorzano A (2009) Mitochondrial dynamics in mammalian health and disease. Physiol Rev 89: 799-845. doi: 10.1152/physrev.00030.2008
|
| [24] |
Alexander C, Votruba M, Pesch U, et al. (2000) OPA1, encoding a dynamin-related GTPase, is mutated in autosomal dominant optic atrophy linked to chromosome 3q28. Nat Genet 26: 211-215. doi: 10.1038/79944
|
| [25] |
Detmer S, Chan D (2007) Functions and dysfunctions of mitochondrial dynamics. Nat Rev Mol Cell Biol 8: 870-879. doi: 10.1038/nrm2275
|
| [26] | Ono T, Isobe K, Nakada K, et al. (2001) Human cells are protected from mitochondrial dysfunction by complementation of DNA products in fused mitochondria. Exp Neurol 28: 272-275. |
| [27] |
Delettre C, Lenaers G, Griffoin JM, et al. (2000) Nuclear gene OPA1, encoding a mitochondrial dynamin-related protein, is mutated in dominant optic atrophy. Nat Genet 26: 207-210. doi: 10.1038/79936
|
| [28] |
Hudson G, Amati-Bonneau P, Blakely EL, et al. (2008) Mutation of OPA1 causes dominant optic atrophy with external ophthalmoplegia, ataxia, deafness and multiple mitochondrial DNA deletions: a novel disorder of mtDNA maintenance. Brain 131: 329-337. doi: 10.1093/brain/awm272
|
| [29] |
Uo T, Dworzak J, Kinoshita C, et al. (2009) Drp1 levels constitutively regulate mitochondrial dynamics and cell survival in cortical neurons. Exp Neurol 218: 274-285. doi: 10.1016/j.expneurol.2009.05.010
|
| [30] |
Ishihara N, Nomura M, Jofuku A, et al. (2009) Mitochondrial fission factor Drp1 is essential for embryonic development and synapse formation in mice. Nat Cell Biol 11: 958-966. doi: 10.1038/ncb1907
|
| [31] |
Darshi M, Mendiola VL, Mackey MR, et al. (2011) ChChd3, an inner mitochondrial membrane protein, is essential for maintaining crista integrity and mitochondrial function. J Biol Chem 286: 2918-2932. doi: 10.1074/jbc.M110.171975
|
| [32] |
Koch A, Yoon Y, Bonekamp NA, et al. (2005) A Role for Fis1 in Both Mitochondrial and Peroxisomal Fission in Mammalian Cells. Mol Biol Cell 16: 5077-5086. doi: 10.1091/mbc.e05-02-0159
|
| [33] |
Serasinghe M, Yoon Y (2008) The mitochondrial outer membrane protein hFis1 regulates mitochondrial morphology and fission through self-interaction. Exp Cell Res 314: 3494-3507. doi: 10.1016/j.yexcr.2008.09.009
|
| [34] |
Otera H, Wang C, Cleland MM, et al. (2010) Mff is an essential factor for mitochondrial recruitment of Drp1 during mitochondrial fission in mammalian cells. J Cell Biol 191: 1141-1158. doi: 10.1083/jcb.201007152
|
| [35] |
Knott AB, Perkins G, Schwarzenbacher R, et al. (2008) Mitochondrial fragmentation in neurodegeneration. Nat Rev Neurosci 9: 505-518. doi: 10.1038/nrn2417
|
| [36] |
Niemann A, Ruegg M, Padula VL, et al. (2005) Ganglioside-induced differentiation associated protein 1 is a regulator of the mitochondrial network: new implications for Charcot-Marie-Tooth disease. J Cell Biol 170: 1067-1078. doi: 10.1083/jcb.200507087
|
| [37] |
Wagner KM, Rüegg M, Niemann A, et al. (2009) Targeting and function of the mitochondrial fission factor GDAP1 are dependent on its tail-anchor. PloS One 4: e5160. doi: 10.1371/journal.pone.0005160
|
| [38] |
Chen H, Detmer SA, Ewald AJ, et al. (2003) Mitofusins Mfn1 and Mfn2 coordinately regulate mitochondrial fusion and are essential for embryonic development. J Cell Biol 160: 189-200. doi: 10.1083/jcb.200211046
|
| [39] |
Wakabayashi J, Zhang Z, Wakabayashi N, et al. (2009) The dynamin-related GTPase Drp1 is required for embryonic and brain development in mice. J Cell Biol 186: 805-816. doi: 10.1083/jcb.200903065
|
| [40] |
Chu CT (2019) Mechanisms of selective autophagy and mitophagy: Implications for neurodegenerative diseases. Neurobiol Dis 122: 23-34. doi: 10.1016/j.nbd.2018.07.015
|
| [41] |
Kageyama Y, Zhang Z, Roda R, et al. (2012) Mitochondrial division ensures the survival of postmitotic neurons by suppressing oxidative damage. J Cell Biol 197: 535-551. doi: 10.1083/jcb.201110034
|
| [42] |
Lutz AK, Exner N, Fett ME, et al. (2009) Loss of parkin or PINK1 function increases Drp1-dependent mitochondrial fragmentation. J Biol Chem 284: 22938-22951. doi: 10.1074/jbc.M109.035774
|
| [43] |
Matsuda N, Sato S, Shiba K, et al. (2010) PINK1 stabilized by mitochondrial depolarization recruits Parkin to damaged mitochondria and activates latent Parkin for mitophagy. J Cell Biol 189: 211-221. doi: 10.1083/jcb.200910140
|
| [44] |
Narendra DP, Jin SM, Tanaka A, et al. (2010) PINK1 is selectively stabilized on impaired mitochondria to activate Parkin. PLoS Biol 8: e1000298. doi: 10.1371/journal.pbio.1000298
|
| [45] |
Zhu J, Dagda RK, CHu CT (2011) Monitoring mitophagy in neuronal cell cultures. Methods Mol Biol 793: 325-339. doi: 10.1007/978-1-61779-328-8_21
|
| [46] |
Knott AB, Bossy-Wetzel E (2009) Impairing the Mitochondrial Fission and Fusion Balance: A New Mechanism of Neurodegeneration. Ann N Y Acad Sci 1147: 283-292. doi: 10.1196/annals.1427.030
|
| [47] |
Martin LJ (2010) Mitochondrial and Cell Death Mechanisms in Neurodegenerative Diseases. Pharmaceuticals 3: 839-915. doi: 10.3390/ph3040839
|
| [48] |
Corrado M, Scorrano L, Campello S (2012) Mitochondrial Dynamics in Cancer and Neurodegenerative and Neuroinflammatory Disease. Int J Cell Biol 2012. doi: 10.1155/2012/729290
|
| [49] |
Su B, Wang X, Zheng L, et al. (2010) Abnormal mitochondrial dynamics and neurodegenerative diseases. Biochim Biophys Acta 1802: 135-142. doi: 10.1016/j.bbadis.2009.09.013
|
| [50] |
Dauer W, Przedborski S (2003) Parkinson's disease: mechanisms and models. Neuron 39: 889-909. doi: 10.1016/S0896-6273(03)00568-3
|
| [51] |
Chang D, Nalls MA, Hallgrmsdóttir IB, et al. (2017) A meta-analysis of genome-wide association studies identifies 17 new Parkinson's disease risk loci. Nat Genet 49: 1511. doi: 10.1038/ng.3955
|
| [52] |
Olanow C, Tatton W (1999) Etiology and pathogenesis of Parkinson's disease. Annu Rev Neurosci 22: 123-144. doi: 10.1146/annurev.neuro.22.1.123
|
| [53] |
Koh H, Chung J (2010) PINK1 and Parkin to control mitochondria remodeling. Anat Cell Biol 43: 179-184. doi: 10.5115/acb.2010.43.3.179
|
| [54] |
Narendra D, Tanaka A, Suen DF, et al. (2008) Parkin is recruited selectively to impaired mitochondria and promotes their autophagy. J Cell Biol 183: 795-803. doi: 10.1083/jcb.200809125
|
| [55] |
Billingsley KJ, Barbosa IA, Bandrés-Ciga S, et al. (2019) Mitochondria function associated genes contribute to Parkinson's Disease risk and later age at onset. NPJ Parkinson's Dis 5: 8. doi: 10.1038/s41531-019-0080-x
|
| [56] |
Panayiotou C, Solaroli N, Johansson M, et al. (2010) Evidence of an intact N-terminal translocation sequence of human mitochondrial adenylate kinase 4. Int J Biochem Cell Biol 42: 62-69. doi: 10.1016/j.biocel.2009.09.007
|
| [57] |
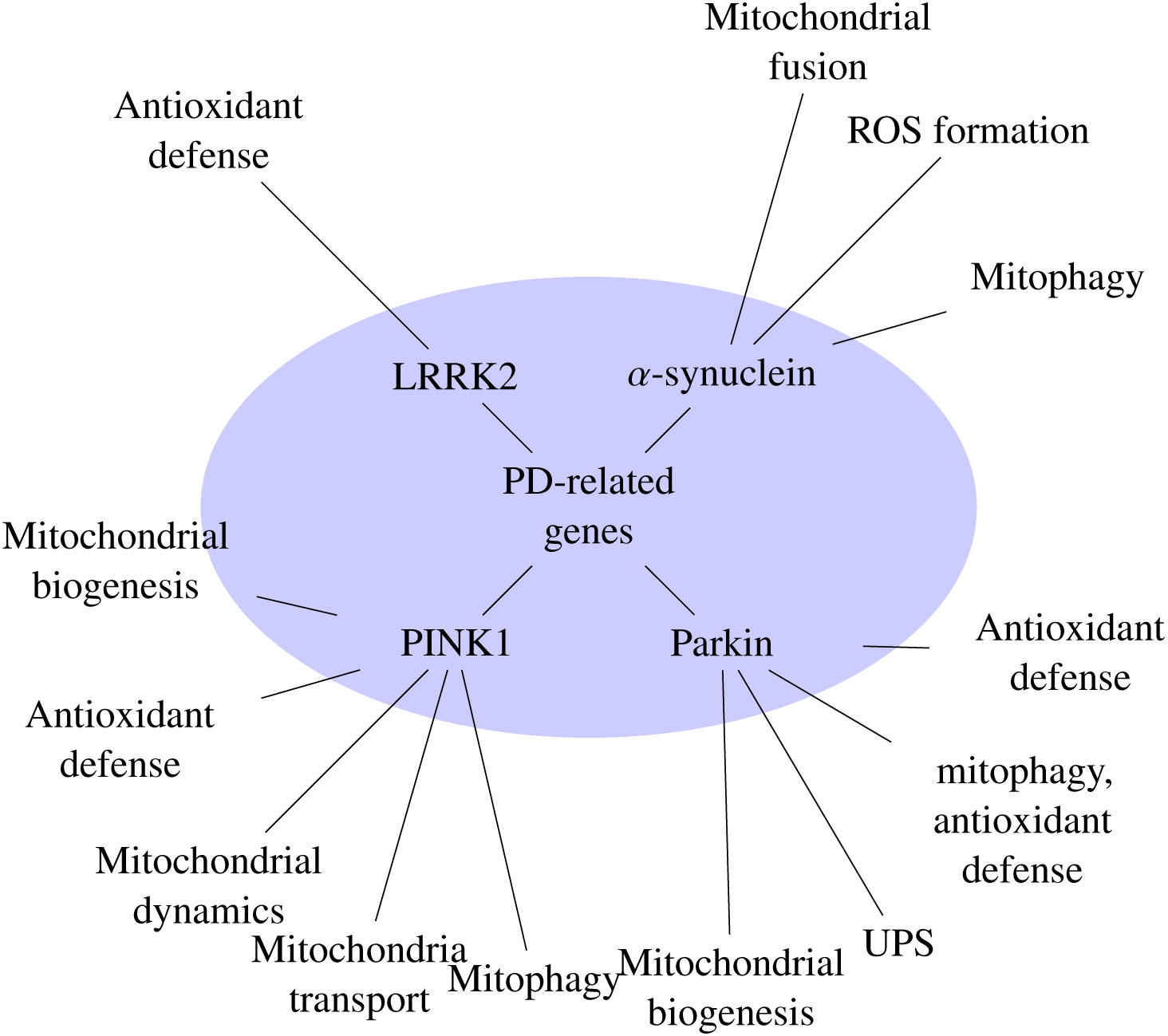
Sai Y, Zou Z, Peng K, et al. (2012) The Parkinson's disease-related genes act in mitochondrial homeostasis. Neurosci Biobehav Rev 36: 2034-2043. doi: 10.1016/j.neubiorev.2012.06.007
|
| [58] |
Perier C, Vila M (2012) Mitochondrial biology and Parkinson's disease. Cold Spring Harb Perspect Med 2: a009332. doi: 10.1101/cshperspect.a009332
|
| [59] |
Comellas G, Lemkau LR, Nieuwkoop AJ, et al. (2011) Structured regions of α-synuclein fibrils include the early-onset Parkinson's disease mutation sites. J Mol Biol 411: 881-895. doi: 10.1016/j.jmb.2011.06.026
|
| [60] |
Ulmer TS, Bax A (2005) Comparison of structure and dynamics of micelle-bound human α-synuclein and Parkinson disease variants. J Biol Chem 280: 43179-43187. doi: 10.1074/jbc.M507624200
|
| [61] |
Nakamura K, Nemani VM, Azarbal F, et al. (2011) Direct membrane association drives mitochondrial fission by the Parkinson disease-associated protein α-synuclein. J Biol Chem 286: 20710-20726. doi: 10.1074/jbc.M110.213538
|
| [62] |
Mori H, Kondo T, Yokochi M, et al. (1998) Pathologic and biochemical studies of juvenile parkinsonism linked to chromosome 6q. Neurology 51: 890-892. doi: 10.1212/WNL.51.3.890
|
| [63] |
Mizuno Y, Hattori N, Yoshino H, et al. (2006) Progress in familial Parkinson's disease. Parkinson's Disease and Related Disorders Springer, 191-204. doi: 10.1007/978-3-211-45295-0_30
|
| [64] |
Lodi R, Tonon C, Valentino M, et al. (2004) Deficit of in vivo mitochondrial ATP production in OPA1-related dominant optic atrophy. Ann Neurol 56: 719-723. doi: 10.1002/ana.20278
|
| [65] |
Mayorov V, Lowrey A, Biousse V, et al. (2008) Mitochondrial oxidative phosphorylation in autosomal dominant optic atrophy. BMC Biochem 9: 22. doi: 10.1186/1471-2091-9-22
|
| [66] |
Zanna C, Ghelli A, Porcelli AM, et al. (2008) OPA1 mutations associated with dominant optic atrophy impair oxidative phosphorylation and mitochondrial fusion. Brain 131: 352-367. doi: 10.1093/brain/awm335
|
| [67] | Krajewski K, Lewis R, Fuerst D, et al. (2000) Neurological dysfunction and axonal degeneration in Charcot-Marie-Tooth disease type 1A. Brain 44: 1299-1304. |
| [68] |
Berger P, Young P, Suter U (2002) Molecular cell biology of Charcot-Marie-Tooth disease. Neurogenetics 4: 1-15. doi: 10.1007/s10048-002-0130-z
|
| [69] |
Baloh R, Schmidt R, Pestronk A, et al. (2007) Altered axonal mitochondrial transport in the pathogenesis of Charcot-Marie-Tooth disease from mitofusin 2 mutations. J Neurosci 27: 422-430. doi: 10.1523/JNEUROSCI.4798-06.2007
|
| [70] |
Züchner S, Vance J (2006) Molecular genetics of autosomal-dominant axonal Charcot-Marie-Tooth disease. Neuromolecular Med 8: 63-74. doi: 10.1385/NMM:8:1-2:63
|
| [71] | Sme F (2010) MFN2 mutations cause severe phenotypes in most patients with CMT2A. Neurology 125: 245-256. |
| [72] |
Cerveny K, Tamura Y (2007) Regulation of mitochondrial fusion and division. Trends Cell Biol 17: 563-569. doi: 10.1016/j.tcb.2007.08.006
|
| [73] |
Poole AC, Thomas RE, Andrews LA, et al. (2008) The PINK1/Parkin pathway regulates mitochondrial morphology. Proc Natl Acad Sci U S A 105: 1638-1643. doi: 10.1073/pnas.0709336105
|
| [74] |
Yang Y, Ouyang Y, Yang L, et al. (2008) Pink1 regulates mitochondrial dynamics through interaction with the fission/fusion machinery. Proc Natl Acad Sci U S A 105: 7070-7075. doi: 10.1073/pnas.0711845105
|
| [75] |
Valente EM, Abou-Sleiman PM, Caputo V, et al. (2004) Hereditary early-onset Parkinson's disease caused by mutations in PINK1. Science 304: 1158-1160. doi: 10.1126/science.1096284
|
| [76] |
Rothfuss O, Fischer H, Hasegawa T, et al. (2009) Parkin protects mitochondrial genome integrity and supports mitochondrial DNA repair. Hum Mol Genet 18: 3832-3850. doi: 10.1093/hmg/ddp327
|
| [77] |
Mortiboys H, Thomas KJ, Koopman WJ, et al. (2008) Mitochondrial function and morphology are impaired in parkin-mutant fibroblasts. Ann Neurol 64: 555-565. doi: 10.1002/ana.21492
|
| [78] |
Büttner S, Bitto A, Ring J, et al. (2008) Functional mitochondria are required for α-synuclein toxicity in aging yeast. J Biol Chem 283: 7554-7560. doi: 10.1074/jbc.M708477200
|
| [79] |
Banerjee K, Sinha M, Pham CLL, et al. (2010) α-Synuclein induced membrane depolarization and loss of phosphorylation capacity of isolated rat brain mitochondria: Implications in Parkinson's disease. FEBS Lett 584: 1571-1576. doi: 10.1016/j.febslet.2010.03.012
|
| [80] |
Yao C, El Khoury R, Wang W, et al. (2010) LRRK2-mediated neurodegeneration and dysfunction of dopaminergic neurons in a Caenorhabditis elegans model of Parkinson's disease. Neurobiol Dis 40: 73-81. doi: 10.1016/j.nbd.2010.04.002
|
| [81] |
Wang X, Yan MH, Fujioka H, et al. (2012) LRRK2 regulates mitochondrial dynamics and function through direct interaction with DLP1. Hum Mol Genet 21: 1931-1944. doi: 10.1093/hmg/dds003
|
| [82] |
Pich S, Bach D, Briones P, et al. (2005) The Charcot–Marie–Tooth type 2A gene product, Mfn2, up-regulates fuel oxidation through expression of OXPHOS system. Hum Mol Genet 14: 1405-1415. doi: 10.1093/hmg/ddi149
|
| [83] |
Detmer SA, Chan DC (2007) Complementation between mouse Mfn1 and Mfn2 protects mitochondrial fusion defects caused by CMT2A disease mutations. J Cell Biol 176: 405-414. doi: 10.1083/jcb.200611080
|
| [84] |
Niemann A, Berger P, Suter U (2006) Pathomechanisms of Mutant Proteins in Charcot-Marie-Tooth Disease. NeuroMolecular Med 8: 217-242. doi: 10.1385/NMM:8:1-2:217
|
| [85] |
Noack R, Frede S, Albrecht P, et al. (2012) Charcot–Marie–Tooth disease CMT4A: GDAP1 increases cellular glutathione and the mitochondrial membrane potential. Hum Mol Genet 21: 150-162. doi: 10.1093/hmg/ddr450
|
| [86] |
Cho D, Nakamura T, Fang J, et al. (2009) S-nitrosylation of Drp1 mediates beta-amyloid-related mitochondrial fission and neuronal injury. Science 324: 102-105. doi: 10.1126/science.1171091
|
| [87] |
Olichon A, Baricault L, Gas N (2003) Loss of OPA1 perturbates the mitochondrial inner membrane structure and integrity, leading to cytochrome c release and apoptosis. J Biol Chem 278: 7743-7746. doi: 10.1074/jbc.C200677200
|
| [88] |
Lin MT, Beal MF (2006) Mitochondrial dysfunction and oxidative stress in neurodegenerative diseases. Nature 443: 787. doi: 10.1038/nature05292
|
| [89] |
Wang X, Wang W, Li L, et al. (2014) Oxidative stress and mitochondrial dysfunction in Alzheimer's disease. Biochim Biophys Acta 1842: 1240-1247. doi: 10.1016/j.bbadis.2013.10.015
|
| [90] |
Smith MA, Rottkamp CA, Nunomura A, et al. (2000) Oxidative stress in Alzheimer's disease. Biochim Biophys Acta 1502: 139-144. doi: 10.1016/S0925-4439(00)00040-5
|
| [91] |
Hirai K, Aliev G, Nunomura A, et al. (2001) Mitochondrial abnormalities in Alzheimer's disease. J Neurosci 21: 3017-3023. doi: 10.1523/JNEUROSCI.21-09-03017.2001
|
| [92] |
Sun A, Chen Y (1998) Oxidative stress and neurodegenerative disorders. J Biomed Sci 5: 401-414. doi: 10.1007/BF02255928
|
| [93] |
Islam MT (2017) Oxidative stress and mitochondrial dysfunction-linked neurodegenerative disorders. Neurol Res 39: 73-82. doi: 10.1080/01616412.2016.1251711
|
| [94] |
Wang X, Su B, Fujioka H, et al. (2008) Dynamin-like protein 1 reduction underlies mitochondrial morphology and distribution abnormalities in fibroblasts from sporadic Alzheimer's disease patients. Am J Pathol 173: 470-482. doi: 10.2353/ajpath.2008.071208
|
| [95] |
Wang X, Su B, Siedlak SL, et al. (2008) Amyloid-β overproduction causes abnormal mitochondrial dynamics via differential modulation of mitochondrial fission/fusion proteins. Proc Natl Acad Sci 105: 19318-19323. doi: 10.1073/pnas.0804871105
|
| [96] |
Martin M, Hurd D, Saxton W (1999) Kinesins in the nervous system. Cell Mol Life Sci 56: 200-216. doi: 10.1007/s000180050422
|
| [97] |
Schnapp BJ, Reese TS (1989) Dynein is the motor for retrograde axonal transport of organelles. Proc Natl Acad Sci U S A 86: 1548-1552. doi: 10.1073/pnas.86.5.1548
|
| [98] | Tabb JS, Molyneaux BJ, Cohen DL, et al. (1998) Transport of ER vesicles on actin filaments in neurons by myosin V. J Cell Sci 111: 3221-3234. |
| [99] |
Ligon LA, Steward O (2000) Role of microtubules and actin filaments in the movement of mitochondria in the axons and dendrites of culture hippocampal neurons. Journal Comp Neur 427: 351-361. doi: 10.1002/1096-9861(20001120)427:3<351::AID-CNE3>3.0.CO;2-R
|
| [100] |
Hirokawa N (1998) Kinesin and dynein superfamily proteins and the mechanism of organelle transport. Science 279: 519-526. doi: 10.1126/science.279.5350.519
|
| [101] |
Gross SP (2004) Hither and yon: A review of bi-directional microtubule-based transport. Phys Biol 1: R1-R11. doi: 10.1088/1478-3967/1/2/R01
|
| [102] |
Ligon LA, Tokito M, Finklestein JM, et al. (2004) A direct interaction between cytoplasmic dynein and kinesin I may coordinate motor activity. J Biol Chem 279: 19201-19208. doi: 10.1074/jbc.M313472200
|
| [103] |
Stowers R, Megeath L, Górska-Andrzejak J, et al. (2002) Axonal transport of mitochondria to synapses depends on milton, a novel Drosophila protein. Neuron 36: 1063-1077. doi: 10.1016/S0896-6273(02)01094-2
|
| [104] |
Guo X, Macleod G, Wellington A, et al. (2005) The GTPase dMiro is required for axonal transport of mitochondria to Drosophila synapses. Neuron 47: 379-393. doi: 10.1016/j.neuron.2005.06.027
|
| [105] |
Russo G, Louie K, Wellington A, et al. (2009) Drosophila Miro is required for both anterograde and retrograde axonal mitochondrial transport. J Neurosci 29: 5443-5455. doi: 10.1523/JNEUROSCI.5417-08.2009
|
| [106] |
Koutsopoulos OS, Laine D, Osellame L, et al. (2010) Human miltons associate with mitochondria and induce microtubule-dependent remodeling of mitochondrial networks. Biochim Biophys Acta 1803: 564-574. doi: 10.1016/j.bbamcr.2010.03.006
|
| [107] |
Wang X, Winter D, Ashrafi G, et al. (2011) PINK1 and Parkin target Miro for phosphorylation and degradation to arrest mitochondrial motility. Cell 147: 893-906. doi: 10.1016/j.cell.2011.10.018
|
| [108] |
Hsieh CH, Shaltouki A, Gonzalez AE, et al. (2016) Functional impairment in miro degradation and mitophagy is a shared feature in familial and sporadic Parkinson's disease. Cell Stem Cell 19: 709-724. doi: 10.1016/j.stem.2016.08.002
|
| [109] |
Baloh RH (2008) Mitochondrial dynamics and peripheral neuropathy. Neuroscientist 14: 12-18. doi: 10.1177/1073858407307354
|
| [110] |
Misko A, Jiang S, Wegorzewska I, et al. (2010) Mitofusin 2 is necessary for transport of axonal mitochondria and interacts with the Miro/Milton complex. J Neurosci 30: 4232-4240. doi: 10.1523/JNEUROSCI.6248-09.2010
|
| [111] |
Ikenaka K, Katsuno M, Kawai K, et al. (2012) Disruption of axonal transport in motor neuron diseases. Int J Mol Sci 13: 1225-1238. doi: 10.3390/ijms13011225
|
| [112] |
Mandal A, Drerup CM (2019) Axonal transport and mitochondrial function in neurons. Front Cell Neurosci 13: 373. doi: 10.3389/fncel.2019.00373
|
| [113] |
Ader NR (2016) Seeking an in vivo neuronal context for the PINK1/Parkin pathway. J Neurosci 36: 11165-11167. doi: 10.1523/JNEUROSCI.2525-16.2016
|
| [114] |
Metaxakis A, Ploumi C, Tavernarakis N (2018) Autophagy in age-associated neurodegeneration. Cells 7: 37. doi: 10.3390/cells7050037
|
| [115] |
Rintoul GL, Reynolds IJ (2010) Mitochondrial trafficking and morphology in neuronal injury. Biochim Biophys Acta 1802: 143-150. doi: 10.1016/j.bbadis.2009.09.005
|
| [116] |
Chang DT, Reynolds IJ (2006) Mitochondrial trafficking and morphology in healthy and injured neurons. Prog Neurobiol 80: 241-268. doi: 10.1016/j.pneurobio.2006.09.003
|
| [117] |
Weihofen A, Thomas K, Ostaszewski B, et al. (2009) Pink1 forms a multiprotein complex with miro and milton, linking Pink1 function to mitochondrial trafficking. Biochemistry 48: 2045-2052. doi: 10.1021/bi8019178
|
| [118] |
Yang F, Jiang Q, Zhao J, et al. (2005) Parkin stabilizes microtubules through strong binding mediated by three independent domains. J Biol Chem 280: 17154-17162. doi: 10.1074/jbc.M500843200
|
| [119] |
Lee H, Khoshaghideh F, Lee S, et al. (2006) Impairment of microtubule-dependent trafficking by overexpression of α-synuclein. European J Neurosci 24: 3153-3162. doi: 10.1111/j.1460-9568.2006.05210.x
|
| [120] |
Gillardon F (2009) Leucine-rich repeat kinase 2 phosphorylates brain tubulin-beta isoforms and modulates microtubule stability—A point of convergence in Parkinsonian neurodegeneration? J Neurochem 10: 1514-1522. doi: 10.1111/j.1471-4159.2009.06235.x
|
| [121] |
Braak E, Sandmann-Keil D, Rüb U, et al. (2001) α-Synuclein immunopositive Parkinson's disease-related inclusion bodies in lower brain stem nuclei. Acta Neuropathol 101: 195-201. doi: 10.1007/s004010000247
|
| [122] |
Liu S, Sawada T, Lee S, et al. (2012) Parkinson's disease–associated kinase PINK1 regulates Miro protein level and axonal transport of mitochondria. PLoS Genet 8: e1002537. doi: 10.1371/journal.pgen.1002537
|
| [123] |
Chu Y, Morfini GA, Langhamer LB, et al. (2012) Alterations in axonal transport motor proteins in sporadic and experimental Parkinson's disease. Brain 135: 2058-2073. doi: 10.1093/brain/aws133
|
| [124] |
Abeliovich A, Gitler AD (2016) Defects in trafficking bridge Parkinson's disease pathology and genetics. Nature 539: 207. doi: 10.1038/nature20414
|
| [125] |
Züchner S, Mersiyanova I, Muglia M, et al. (2004) Mutations in the mitochondrial GTPase mitofusin 2 cause Charcot-Marie-Tooth neuropathy type 2A. Nat Genet 36: 449-451. doi: 10.1038/ng1341
|
| [126] |
Patzkó A, Shy ME (2011) Update on Charcot-Marie-Tooth Disease. Curr Neurol Neurosci Rep 11: 78-88. doi: 10.1007/s11910-010-0158-7
|
| [127] |
Shy M (2004) Charcot-Marie-Tooth disease: An update. Curr Opin Neurol 17: 579-585. doi: 10.1097/00019052-200410000-00008
|
| [128] |
Loiseau D, Chevrollier A, Verny C, et al. (2007) Mitochondrial coupling defect in Charcot–Marie–Tooth type 2A disease. Ann Neurol 61: 315-323. doi: 10.1002/ana.21086
|
| [129] |
Stokin G, Lillo C, Falzone T, et al. (2005) Axonopathy and transport deficits early in the pathogenesis of Alzheimer's diseases. Science 307: 1282-1288. doi: 10.1126/science.1105681
|
| [130] |
Wang X, Su B, Lee H (2009) Impaired balance of mitochondrial fission and fusion in Alzheimer's disease. J Neurosci 29: 9090-9103. doi: 10.1523/JNEUROSCI.1357-09.2009
|
| [131] |
Vickers JC, King AE, Woodhouse A, et al. (2009) Axonopathy and cytoskeletal disruption in degenerative diseases of the central nervous system. Brain Res Bull 80: 217-223. doi: 10.1016/j.brainresbull.2009.08.004
|
| [132] |
De Vos K, Chapman A, Tennant M, et al. (2007) Familial amyotrophic lateral sclerosis-linked SOD1 mutants perturb fast axonal transport to reduce axonal mitochondria content. Hum Mol Genet 16: 2720-2728. doi: 10.1093/hmg/ddm226
|
| [133] |
Shi P, Ström A, Gal J, et al. (2010) Effects of ALS-related SOD1 mutants on dynein- and KIF5-mediated retrograde and anterograde axonal transport. Biochim Biophys Acta 1802: 707-716. doi: 10.1016/j.bbadis.2010.05.008
|
| [134] |
Chang D, Rintoul G, Pandipati S, et al. (2006) Mutant huntingtin aggregates impair mitochondrial movement and trafficking in cortical neurons. Neurobiol Dis 22: 388-400. doi: 10.1016/j.nbd.2005.12.007
|
| [135] |
Trushina E, Dyer R, Badger J, et al. (2004) Mutant huntingtin impairs axonal trafficking in mammalian neurons in vivo and in vitro. Mol Cell Biol 24: 8195-8209. doi: 10.1128/MCB.24.18.8195-8209.2004
|
| [136] |
Bach D, Pich S, Soriano FX (2003) Mitofusin-2 determines mitochondrial network architecture and mitochondrial metabolism: a novel regulatory mechanism altered in obesity. J Biol Chem 278: 17190-17197. doi: 10.1074/jbc.M212754200
|
| [137] |
Chen H, Chomyn A, Chan DC (2005) Disruption of fusion results in mitochondrial heterogeneity and dysfunction. J Biol Chem 280: 26185-26192. doi: 10.1074/jbc.M503062200
|
| [138] |
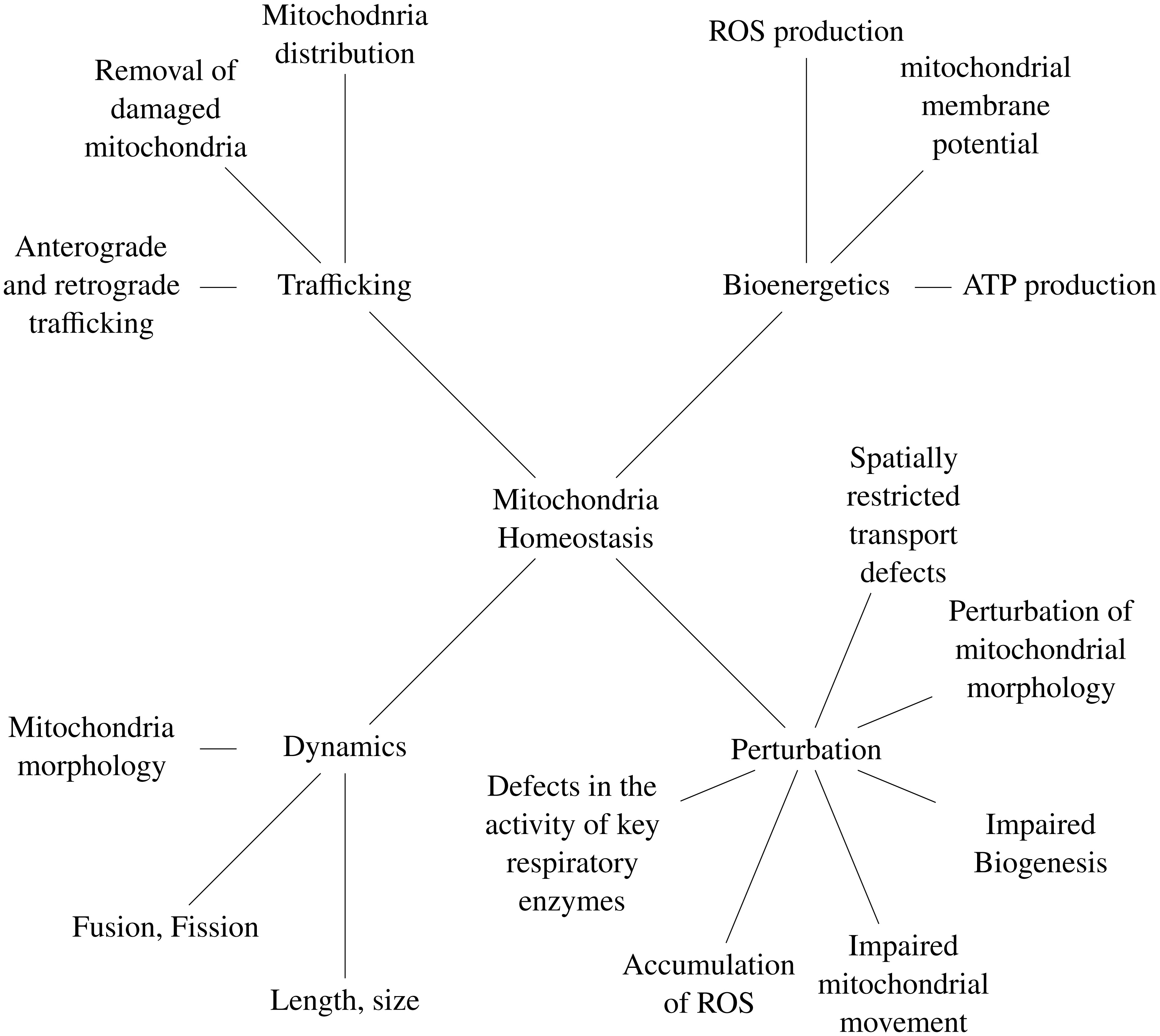
Van Laar VS, Berman SB (2013) The interplay of neuronal mitochondrial dynamics and bioenergetics: implications for Parkinson's disease. Neurobiol Dis 51: 43-55. doi: 10.1016/j.nbd.2012.05.015
|
| [139] |
Coskun P, Wyrembak J, Schriner SE, et al. (2012) A mitochondrial etiology of Alzheimer and Parkinson disease. Biochim Biophys Acta 1820: 553-564. doi: 10.1016/j.bbagen.2011.08.008
|
| [140] |
Ryan BJ, Hoek S, Fon EA, et al. (2015) Mitochondrial dysfunction and mitophagy in Parkinson's: from familial to sporadic disease. Trends Biochem Sci 40: 200-210. doi: 10.1016/j.tibs.2015.02.003
|
| [141] |
Van Laar VS, Berman SB (2009) Mitochondrial dynamics in Parkinson's disease. Exp Neurol 218: 247-256. doi: 10.1016/j.expneurol.2009.03.019
|
| [142] |
Requejo-Aguilar R, Bolaños JP (2016) Mitochondrial control of cell bioenergetics in Parkinson's disease. Free Radic Biol Med 100: 123-137. doi: 10.1016/j.freeradbiomed.2016.04.012
|
| [143] |
Abramov AY, Gegg M, Grunewald A, et al. (2011) Bioenergetic consequences of PINK1 mutations in Parkinson disease. PLoS One 6: e25622. doi: 10.1371/journal.pone.0025622
|
| [144] |
Lansbury PT, Lashuel HA (2006) A century-old debate on protein aggregation and neurodegeneration enters the clinic. Nature 443: 774-779. doi: 10.1038/nature05290
|
| [145] |
Jain S, Wood NW, Healy DG (2005) Molecular genetic pathways in Parkinson's disease: A review. Clin Sci 109: 355-364. doi: 10.1042/CS20050106
|
| [146] |
Winklhofer KF, Haass C (2010) Mitochondrial dysfunction in Parkinson's disease. Biochim Biophy Acta 1802: 29-44. doi: 10.1016/j.bbadis.2009.08.013
|
| [147] |
Chen C, Turnbull DM, Reeve AK (2019) Mitochondrial Dysfunction in Parkinson's Disease—Cause or Consequence? Biology 8: 38. doi: 10.3390/biology8020038
|
| [148] |
Diot A, Morten K, Poulton J (2016) Mitophagy plays a central role in mitochondrial ageing. Mamm Genome 27: 381-395. doi: 10.1007/s00335-016-9651-x
|
| [149] |
Noda S, Sato S, Fukuda T, et al. (2019) Loss of Parkin contributes to mitochondrial turnover and dopaminergic neuronal loss in aged mice. Neurobiol Dis 136: 104717. doi: 10.1016/j.nbd.2019.104717
|
| [150] |
Fivenson EM, Lautrup S, Sun N, et al. (2017) Mitophagy in neurodegeneration and aging. Neurochem Int 109: 202-209. doi: 10.1016/j.neuint.2017.02.007
|
| [151] |
Ebrahimi-Fakhari D, Wahlster L, McLean PJ (2012) Protein degradation pathways in Parkinson's disease: curse or blessing. Acta Neuropathol 124: 153-172. doi: 10.1007/s00401-012-1004-6
|
| [152] |
Wang W, Wang X, Fujioka H, et al. (2016) Parkinson's disease–associated mutant VPS35 causes mitochondrial dysfunction by recycling DLP1 complexes. Nat Med 22: 54. doi: 10.1038/nm.3983
|
| [153] |
Singleton A, Farrer M, Johnson J, et al. (2003) α-Synuclein locus triplication causes Parkinson's disease. Science 302: 841-841. doi: 10.1126/science.1090278
|
| [154] | Regev A, Silverman W, Shapiro E (2001) Representation and simulation of biochemical processes using the π-calculus process algebra. Pac Symp Biocomput 459-470. |
| [155] | Bonzanni N, Feenstra KA, Fokkink W, et al. (2009) What can formal methods bring to systems biology? International Symposium on Formal Methods Springer, 16-22. |
| [156] |
Theocharopoulou G, Giannakis K, Andronikos T (2015) The mechanism of splitting mitochondria in terms of membrane automata. 2015 IEEE International Symposium on Signal Processing and Information Technology (ISSPIT) IEEE, 397-402. doi: 10.1109/ISSPIT.2015.7394367
|
| [157] |
Theocharopoulou G, Vlamos P (2015) Modeling protein misfolding in Charcot–Marie–Tooth disease. GeNeDis 2014 Springer, 91-102. doi: 10.1007/978-3-319-09012-2_7
|
| [158] |
Theocharopoulou G, Bobori C, Vlamos P (2017) Formal models of biological systems. GeNeDis 2016 Springer, 325-338. doi: 10.1007/978-3-319-56246-9_27
|
| [159] |
Corona JC, Duchen MR (2015) Impaired mitochondrial homeostasis and neurodegeneration: towards new therapeutic targets? J Bioenerg Biomembr 47: 89-99. doi: 10.1007/s10863-014-9576-6
|
| [160] |
Kitada T, Asakawa S, Hattori N, et al. (1998) Mutations in the parkin gene cause autosomal recessive juvenile parkinsonism. Nature 392: 605. doi: 10.1038/33416
|
| [161] |
Healy DG, Falchi M, O'Sullivan SS, et al. (2008) Phenotype, genotype, and worldwide genetic penetrance of LRRK2-associated Parkinson's disease: a case-control study. Lancet Neurol 7: 583-590. doi: 10.1016/S1474-4422(08)70117-0
|
| [162] |
Peng JY, Lin CC, Chen YJ, et al. (2011) Automatic morphological subtyping reveals new roles of caspases in mitochondrial dynamics. PLoS Comput Biol 7: e1002212. doi: 10.1371/journal.pcbi.1002212
|
| [163] |
Tronstad KJ, Nooteboom M, Nilsson LI, et al. (2014) Regulation and quantification of cellular mitochondrial morphology and content. Curr Pharm Des 20: 5634-5652. doi: 10.2174/1381612820666140305230546
|
| [164] |
Zamponi N, Zamponi E, Cannas SA, et al. (2018) Mitochondrial network complexity emerges from fission/fusion dynamics. Sci Rep 8: 363. doi: 10.1038/s41598-017-18351-5
|
| [165] |
Shah SI, Paine JG, Perez C, et al. (2019) Mitochondrial fragmentation and network architecture in degenerative diseases. PloS One 14: e0223014. doi: 10.1371/journal.pone.0223014
|
| [166] |
Toglia P, Demuro A, Mak DOD, et al. (2018) Data-driven modeling of mitochondrial dysfunction in Alzheimer's disease. Cell Calcium 76: 23-35. doi: 10.1016/j.ceca.2018.09.003
|
| [167] |
Dukes AA, Bai Q, Van Laar VS, et al. (2016) Live imaging of mitochondrial dynamics in CNS dopaminergic neurons in vivo demonstrates early reversal of mitochondrial transport following MPP+ exposure. Neurobiol Dis 95: 238-249. doi: 10.1016/j.nbd.2016.07.020
|
| [168] | Alexiou A, Vlamos P (2012) Evidence for early identification of Alzheimer's disease. In arXiv preprint arXiv 1209.4223. |
| [169] |
Lundkvist J, Naslund J (2007) Gamma-secretase: A complex target for Alzheimer's disease. Curr Opin Pharmacol 7: 112-118. doi: 10.1016/j.coph.2006.10.002
|
| [170] |
Evin G, Kenche V (2007) BACE inhibitors as potential therapeutics for Alzheimer's disease. Recent Pat CNS Drug Discov 2: 188-199. doi: 10.2174/157488907782411783
|
| [171] |
Alexiou A, Vlamos P, Rekkas J (2011) Modeling the mitochondrial dysfunction in neurogenerative diseases due to high H+ concentration. Bioinformation 6: 173-175. doi: 10.6026/97320630006173
|
| [172] |
Miller KE, Sheetz MP (2004) Axonal mitochondrial transport and potential are correlated. J Cell Sci 117: 2791-2804. doi: 10.1242/jcs.01130
|
| [173] |
Oliveira JMA (2011) Techniques to investigate neuronal mitochondrial function and its pharmacological modulation. Curr Drug Targets 12: 762-773. doi: 10.2174/138945011795528895
|
| [174] |
Twig G, Hyde B, Shirihai OS (2008) Mitochondrial fusion, fission and autophagy as a quality control axis: the bioenergetic view. Biochim Biophys Acta 1777: 1092-1097. doi: 10.1016/j.bbabio.2008.05.001
|
| [175] |
Onyango IG, Dennis J, Khan SM (2016) Mitochondrial dysfunction in Alzheimer's disease and the rationale for bioenergetics based therapies. Aging Dis 7: 201-214. doi: 10.14336/AD.2015.1007
|
Figures(2) / Tables(2)

Georgia Theocharopoulou. The ubiquitous role of mitochondria in Parkinson and other neurodegenerative diseases[J]. AIMS Neuroscience, 2020, 7(1): 43-65. doi: 10.3934/Neuroscience.2020004







 DownLoad:
DownLoad: