Citation: Byron Bernal, Alfredo Ardila, Monica Rosselli. The Network of Brodmanns Area 22 in Lexico-semantic Processing: A Pooling-data Connectivity Study[J]. AIMS Neuroscience, 2016, 3(3): 306-316. doi: 10.3934/Neuroscience.2016.3.306

| [1] | Mirz F, Ovesen T, Ishizu K, et al. (1999) Stimulus-dependent central processing of auditory stimuli: a PET study. Scand Audiol 28: 161–169. |
| [2] | Goel V, Gold B, Kapur S (1998) Neuroanatomical correlates of human reasoning. J Cogn Neurosci 10: 293–302. |
| [3] | McDermott KB, Petersen SE, Watson JM, et al. (2003) A procedure for identifying regions preferentially activated by attention to semantic and phonological relations using functional magnetic resonance imaging. Neuropsychologia 41: 293–303. |
| [4] |
Chou T-L, Booth JR, Bitan T, et al. (2006) Developmental and skill effects on the neural correlates of semantic processing to visually presented words. Hum Brain Mapp 27: 915–924. doi: 10.1002/hbm.20231

|
| [5] |
McIntosh AR (2000) Towards a network theory of cognition. Neural Netw Off J Int Neural Netw Soc 13: 861–870. doi: 10.1016/S0893-6080(00)00059-9
|
| [6] | Biswal BB, Eldreth DA, Motes MA, et al. (2010) Task-dependent individual differences in prefrontal connectivity. Cereb Cortex N Y N 1991 20: 2188–2197. |
| [7] |
Caclin A, Fonlupt P (2006) Effect of initial fMRI data modeling on the connectivity reported between brain areas. NeuroImage 33: 515–521. doi: 10.1016/j.neuroimage.2006.07.019
|
| [8] |
Wu X, Lu J, Chen K, et al. (2009) Multiple neural networks supporting a semantic task: an fMRI study using independent component analysis. NeuroImage 45: 1347–1358. doi: 10.1016/j.neuroimage.2008.12.050
|
| [9] | Laird AR, Eickhoff SB, Rottschy C, et al. (2013) Networks of task co-activations. NeuroImage 80: 505–514. |
| [10] |
Kohn N, Eickhoff SB, Scheller M, et al. (2014) Neural network of cognitive emotion regulation--an ALE meta-analysis and MACM analysis. NeuroImage 87: 345–355. doi: 10.1016/j.neuroimage.2013.11.001
|
| [11] | Robinson JL, Laird AR, Glahn DC, et al. (2010) Metaanalytic connectivity modeling: delineating the functional connectivity of the human amygdala. Hum Brain Mapp 31: 173–184. |
| [12] |
Eickhoff SB, Laird AR, Grefkes C, et al. (2009) Coordinate-based activation likelihood estimation meta-analysis of neuroimaging data: a random-effects approach based on empirical estimates of spatial uncertainty. Hum Brain Mapp 30: 2907–2926. doi: 10.1002/hbm.20718
|
| [13] |
Ardila A. Bernal B, Rosselli M (2014) Participation of the insula in language revisited: A meta-analytic connectivity study. J Neurolinguistics 29: 31–41. doi: 10.1016/j.jneuroling.2014.02.001
|
| [14] |
Catani M, Mesulam MM, Jakobsen E, et al. (2013) A novel frontal pathway underlies verbal fluency in primary progressive aphasia. Brain J Neurol 136: 2619–2628. doi: 10.1093/brain/awt163
|
| [15] |
Martino J, de Lucas EM, Ibá?ez-Plágaro FJ, et al. (2012) Foix-Chavany-Marie syndrome caused by a disconnection between the right pars opercularis of the inferior frontal gyrus and the supplementary motor area. J Neurosurg 117: 844–850. doi: 10.3171/2012.7.JNS12404
|
| [16] | Nathaniel-James DA, Fletcher P, Frith CD (1997) The functional anatomy of verbal initiation and suppression using the Hayling Test. Neuropsychologia 35: 559–566. |
| [17] | Garn CL, Allen MD, Larsen JD (2009) An fMRI study of sex differences in brain activation during object naming. Cortex J Devoted Study Nerv Syst Behav 45: 610–618. |
| [18] |
Kiyosawa M, Inoue C, Kawasaki T, et al. (1996) Functional neuroanatomy of visual object naming: a PET study. Graefes Arch Clin Exp Ophthalmol Albrecht Von Graefes Arch Für Klin Exp Ophthalmol 234: 110–115. doi: 10.1007/BF00695250
|
| [19] |
Whitney C, Weis S, Krings T, et al. (2009) Task-dependent modulations of prefrontal and hippocampal activity during intrinsic word production. J Cogn Neurosci 21: 697–712. doi: 10.1162/jocn.2009.21056
|
| [20] | Friederici AD, Rüschemeyer S-A, Hahne A, et al. (2003) The role of left inferior frontal and superior temporal cortex in sentence comprehension: localizing syntactic and semantic processes. Cereb Cortex N Y N 1991 13: 170–177. |
| [21] |
Correia J, Formisano E, Valente G, et al. (2014) Brain-based translation: fMRI decoding of spoken words in bilinguals reveals language-independent semantic representations in anterior temporal lobe. J Neurosci Off J Soc Neurosci 34: 332–338. doi: 10.1523/JNEUROSCI.1302-13.2014
|
| [22] |
Alba-Ferrara L, Ellison A, Mitchell RL (2012) Decoding emotional prosody: resolving differences in functional neuroanatomy from fMRI and lesion studies using TMS. Brain Stimulat 5: 347–353. doi: 10.1016/j.brs.2011.06.004
|
| [23] | Benson F, Ardila A (1996) Aphasia, A clinical perspective. New York: Oxford University Pres. |
| [24] | Luria A (1976) Basic problems of neurolinguistics. New York: Mouton. |
| [25] |
Abrahams S, Goldstein LH, Simmons A, et al. (2003) Functional magnetic resonance imaging of verbal fluency and confrontation naming using compressed image acquisition to permit overt responses. Hum Brain Mapp 20: 29–40. doi: 10.1002/hbm.10126
|
| [26] |
Friedman L, Kenny JT, Wise AL, et al. (1998) Brain activation during silent word generation evaluated with functional MRI. Brain Lang 64: 231–256. doi: 10.1006/brln.1998.1953
|
| [27] |
Ahmad Z, Balsamo LM, Sachs BC, et al. (2003) Auditory comprehension of language in young children: neural networks identified with fMRI. Neurology 60: 1598–1605. doi: 10.1212/01.WNL.0000059865.32155.86
|
| [28] | Giraud AL, Kell C, Thierfelder C, et al. (2004) Contributions of sensory input, auditory search and verbal comprehension to cortical activity during speech processing. Cereb Cortex N Y N 1991 14: 247–255. |
| [29] |
S?derfeldt B, Ingvar M, R?nnberg J, et al. (1997) Signed and spoken language perception studied by positron emission tomography. Neurology 49: 82–87. doi: 10.1212/WNL.49.1.82
|
| [30] | Ardila A, Bernal B, Rosselli M (2015) Language and visual perception associations: Meta-analytic connectivity modeling of Brodmann area 37. Behav Neurol. |
| [31] | Turken AU, Dronkers NF (2011) The neural architecture of the language comprehension network: converging evidence from lesion and connectivity analyses. Front Syst Neurosci 5: 1. |
| [32] | Fox PT, Ingham RJ, Ingham JC, et al. (2000) Brain correlates of stuttering and syllable production. A PET performance-correlation analysis. Brain J Neurol 123: 1985–2004. |
| [33] |
Shuster LI, Lemieux SK (2005) An fMRI investigation of covertly and overtly produced mono-and multisyllabic words. Brain Lang 93: 20–31. doi: 10.1016/j.bandl.2004.07.007
|
| [34] | Basho S, Palmer ED, Rubio MA, et al. (2007) Effects of generation mode in fMRI adaptations of semantic fluency: paced production and overt speech. Neuropsychologia 45: 1697–1706. |
| [35] | De Carli D, Garreffa G, Colonnese C, et al. (2007) Identification of activated regions during a language task. Magn Reson Imaging 25: 933–938. |
| [36] |
Price CJ, Green DW, von Studnitz R (1999) A functional imaging study of translation and language switching. Brain J Neurol 122: 2221–2235. doi: 10.1093/brain/122.12.2221
|
| [37] | Baciu MV, Rubin C, Décorps MA, et al. (1999) fMRI assessment of hemispheric language dominance using a simple inner speech paradigm. NMR Biomed 12: 293–298. |
| [38] |
Hirsch J, Moreno DR, Kim KH (2001) Interconnected large-scale systems for three fundamental cognitive tasks revealed by functional MRI. J Cogn Neurosci 13: 389–405. doi: 10.1162/08989290151137421
|
| [39] | Price CJ, Wise RJ, Watson JD, et al. (1994) Brain activity during reading. The effects of exposure duration and task. Brain J Neurol 117: 1255–1269. |
| [40] | Inui T, Otsu Y, Tanaka S, et al. (1998) A functional MRI analysis of comprehension processes of Japanese sentences. Neuroreport 9: 3325–3328. |
| [41] |
Allen K, Pereira F, Botvinick M, et al. (2012) Distinguishing grammatical constructions with fMRI pattern analysis. Brain Lang 123: 174–182. doi: 10.1016/j.bandl.2012.08.005
|
| [42] |
Thompson CK, Bonakdarpour B, Fix SC, et al. (2007) Neural correlates of verb argument structure processing. J Cogn Neurosci 19: 1753–1767. doi: 10.1162/jocn.2007.19.11.1753
|
| [43] | Bechtereva NP, Korotkov AD, Pakhomov SV, et al. (2004) PET study of brain maintenance of verbal creative activity. Int J Psychophysiol Off J Int Organ Psychophysiol 53: 11–20. |
| [44] | Sollmann N, Tanigawa N, Tussis L, et al. (2015) Cortical regions involved in semantic processing investigated by repetitive navigated transcranial magnetic stimulation and object naming. Neuropsychologia 70: 185–195. |
| [45] | Chouinard PA, Goodale MA (2010) Category-specific neural processing for naming pictures of animals and naming pictures of tools: an ALE meta-analysis. Neuropsychologia 48: 409–418. |
| [46] | Barrós-Loscertales A, González J, Pulvermüller F, et al. (2012) Reading salt activates gustatory brain regions: fMRI evidence for semantic grounding in a novel sensory modality. Cereb Cortex N Y N 1991 22: 2554–2563. |
| [47] | Palti D, Ben Shachar M, Hendler T, et al. (2007) Neural correlates of semantic and morphological processing of Hebrew nouns and verbs. Hum Brain Mapp 28: 303–314. |
| [48] | Perani D, Cappa SF, Schnur T, et al. (1999) The neural correlates of verb and noun processing. A PET study. Brain J Neurol 122: 2337–2344. |
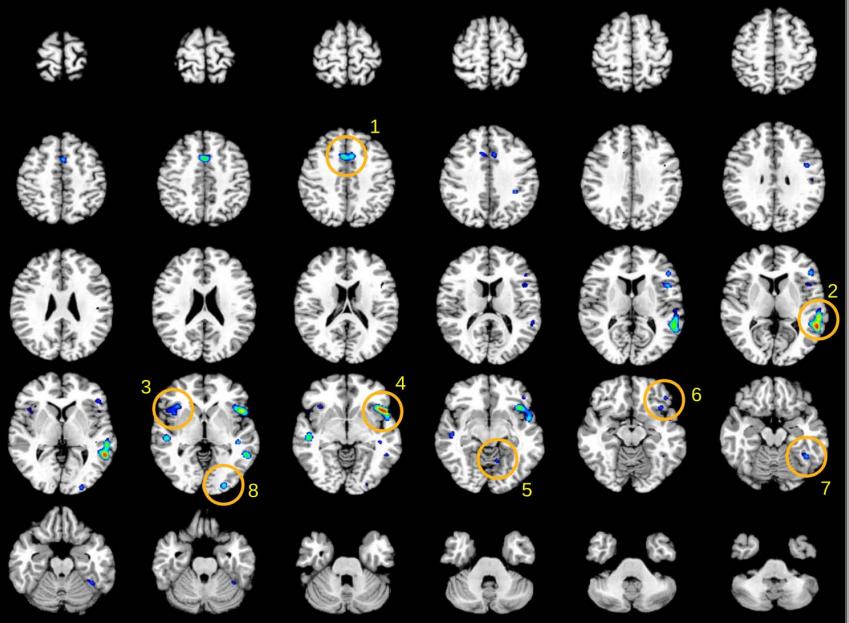
Figures(1) / Tables(1)

Byron Bernal, Alfredo Ardila, Monica Rosselli. The Network of Brodmanns Area 22 in Lexico-semantic Processing: A Pooling-data Connectivity Study[J]. AIMS Neuroscience, 2016, 3(3): 306-316. doi: 10.3934/Neuroscience.2016.3.306







 DownLoad:
DownLoad: