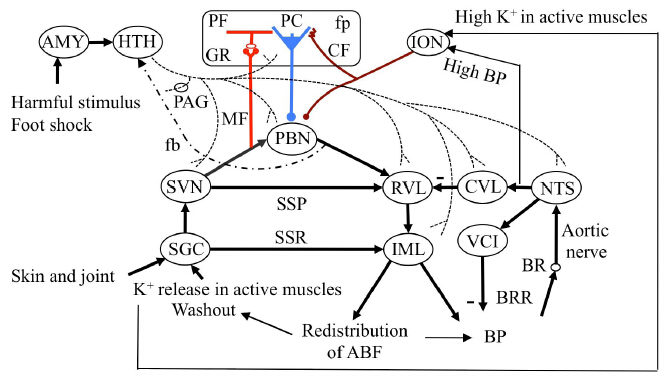
Recent evidence has indicated that, when an animal is exposed to harmful stimuli, hypothalamic orexinergic neurons are activated via the amygdala and in turn tune the neuronal circuits in the spinal cord, brainstem, and an area of the cerebellum (folium-p of the flocculus) by neuromodulation. The animal would then initiate “defense reactions” composed of complex movements and associated cardiovascular responses. To investigate neuronal mechanisms of the defense reactions, Nisimaru et al. (2013) analyzed cardiovascular responses induced by an electric foot shock stimulus to a rabbit and found two major effects. One is redistribution of arterial blood flow from visceral organs to active muscles, and the other is a modest increase in blood pressure. Kainate-induced lesions of folium-p impaired these two effects. Moreover, folium-p Purkinje cells were shown to project to the parabrachial nucleus, one of the major cardiovascular centers in the brainstem. These data indicate that folium-p Purkinje cells regulate cardiovascular defense reactions via parabrachial nucleus under orexin-mediated neuromodulation. In this article, we review these data from the viewpoint that the defense reactions are expressions of “anger and anxiety”, which respectively lead to “fight and flight” behaviors. The present orexin case may provide a model of cerebellohypothalamic interactions via neuropeptides or amines of hypothalamic origin, which may underlie various types of emotion and behavior.
Citation: Masao Ito, Naoko Nisimaru. Cerebellar Control of Defense Reactions under Orexin-mediated Neuromodulation as a Model of Cerebellohypothalamic Interaction[J]. AIMS Neuroscience, 2014, 1(1): 89-95. doi: 10.3934/Neuroscience.2014.1.89
Recent evidence has indicated that, when an animal is exposed to harmful stimuli, hypothalamic orexinergic neurons are activated via the amygdala and in turn tune the neuronal circuits in the spinal cord, brainstem, and an area of the cerebellum (folium-p of the flocculus) by neuromodulation. The animal would then initiate “defense reactions” composed of complex movements and associated cardiovascular responses. To investigate neuronal mechanisms of the defense reactions, Nisimaru et al. (2013) analyzed cardiovascular responses induced by an electric foot shock stimulus to a rabbit and found two major effects. One is redistribution of arterial blood flow from visceral organs to active muscles, and the other is a modest increase in blood pressure. Kainate-induced lesions of folium-p impaired these two effects. Moreover, folium-p Purkinje cells were shown to project to the parabrachial nucleus, one of the major cardiovascular centers in the brainstem. These data indicate that folium-p Purkinje cells regulate cardiovascular defense reactions via parabrachial nucleus under orexin-mediated neuromodulation. In this article, we review these data from the viewpoint that the defense reactions are expressions of “anger and anxiety”, which respectively lead to “fight and flight” behaviors. The present orexin case may provide a model of cerebellohypothalamic interactions via neuropeptides or amines of hypothalamic origin, which may underlie various types of emotion and behavior.

| [1] |
Haines DE, Dietrichs E. (1984) An HRP study of hypothalmo-cerebellar and cerebello-hypothalamic connections in squirrel monkey (Saimiri sciureus). J Comp Neurol 229:559-575. doi: 10.1002/cne.902290409

|
| [2] |
Haines DE, Dietrichs E, Sowa TE. (1984) Hypothalamo-cerebellar and cerebello-hypothalamic pathways: a review and hypothesis concerning cerebellar circuits which may influence autonomic centers affective behavior. Brain Behav Evol 24: 198-220. doi: 10.1159/000121317
|
| [3] |
Bester H, Besson JM, Bernard JF. (1997) Organization of efferent projections from the parabrachial area to the hypothalamus: A Phaseolus vulgaris-leucoagglutinin study in the rat. J Comp Neurol 383: 245-281. doi: 10.1002/(SICI)1096-9861(19970707)383:3<245::AID-CNE1>3.0.CO;2-3
|
| [4] |
Schmahmann JD, Sherman JC. (1998) The cerebellar cognitive affective syndrome. Brain 121:561-579. doi: 10.1093/brain/121.4.561
|
| [5] |
Parvizi J, Anderson SW, Martin CO, et al. (2001) Pathological laughter and crying: a link to the cerebellum. Brain 124: 1708-1719. doi: 10.1093/brain/124.9.1708
|
| [6] |
Andreasen NC, Cohen G, Harris G, et al. (1992) Image processing for the study of brain structure and function: problems and programs. J Neuropsychiat Clinical Neurosci 4: 125-133. doi: 10.1176/jnp.4.2.125
|
| [7] |
Paradiso S, Andreasen NC, Crespo-Facorro B, et al. (2003) Emotions in unmedicated patients with schizophrenia during evaluation with positron emission tomography. Am J Psychiatry 160:1775-1783. doi: 10.1176/appi.ajp.160.10.1775
|
| [8] |
Turner BM, Paradiso S, Marvel CL, et al. (2007) The Cerebellum and Emotional Experience. Neuropsychologia 45: 1331-1341. doi: 10.1016/j.neuropsychologia.2006.09.023
|
| [9] |
Nambu T, Sakurai T, Mizukami K, et al. (1999) Distribution of orexin neurons in the adult rat brain. Brain Res 827: 243-260. doi: 10.1016/S0006-8993(99)01336-0
|
| [10] | Nisimaru N, Mittal C, Shirai Y, et al. (2013) Orexin-neuromodulated cerebellar circuit controls redistribution of arterial blood flows for defense behavior in rabbits. Proc Natl Acad Sci USA 110:14124-14131 |
| [11] |
King JS, Cummings SL, Bishop GA. (1992) Peptides in cerebellar circuit. Prog Neurobiol 39:423-442. doi: 10.1016/0301-0082(92)90014-6
|
| [12] |
Panula P, Takagi H, Inagaki N, et al. (1993) Histamine-containing nerve fibers innervate human cerebellum. Neurosci Lett 160: 53-56. doi: 10.1016/0304-3940(93)90915-8
|
| [13] |
Lind RW, Swanson LW, Ganten D. (1985) Organization of angiotensin II immunoreactive cells and fibers in the rat central nervous system. Neuroendocrinology 40: 2-24. doi: 10.1159/000124046
|
| [14] |
Ito M. (2009) Functional roles of neuropeptides in cerebellar circuits. Neuroscience 162:666-672. doi: 10.1016/j.neuroscience.2009.01.019
|
| [15] |
Marder E, Thirumalai V. (2002) Cellular, synaptic and network effects of neuromodulation. Neural Netw 15: 479-493. doi: 10.1016/S0893-6080(02)00043-6
|
| [16] | Adams DB, Baccelli G, Mancia G, et al. (1969) Cardiovascular changes during naturally elicited fighting behavior in the cat. Am J Physiol 216(5): 1226-1235. |
| [17] | Markgraf CG, Winters RW, Liskowsky DR et al. (1991) Hypothalamic, midbrain and bulbar areas involved in the defense reaction in rabbits. Physiol Behav 49: 493-500. |
| [18] | Silveira MC, Graeff FG (1992) Defense reaction elicited by microinjection of kainic acid into the medial hypothalamus of the rat: Antagonism by a GABAA receptor agonist. Behav Neural Biol57(3): 226-232. |
| [19] |
Kayaba Y, Nakamura A, Kasuya Y, et al. (2003) Attenuated defense response and low basal blood pressure in orexin knockout mice. Am J Physiol Regul Integr Comp Physiol 285: R581-R593. doi: 10.1152/ajpregu.00671.2002
|
| [20] |
Zhang W, Sakurai T, Fukuda Y, et al. (2006) Orexin neuron-mediated skeletal muscle vasodilation and shift of baroreflex during defense response in mice. Am J Physiol Regul Integr Comp Physiol 290: R1654-R1663. doi: 10.1152/ajpregu.00704.2005
|
| [21] | Sakurai T, Amemiya A, Ishii M, et al. (1998) Orexins and orexin receptors: A family of hypothalamic neuropeptides and G protein-coupled receptors that regulate feeding behavior. Cell92(4): 573-585. |
| [22] | Kerman IA, Yates BJ. (1999) Patterning of somatosympathetic reflexes. Am J Physiol 277: R716-R724. |
| [23] | Sato A, Schmidt RF. (1973) Somatosympathetic reflexes: Afferent fibers, central pathways, discharge characteristics. Physiol Rev 53: 916-947. |
| [24] | Eriksson KS, Sergeeva O, Brown RE, et al. (2001) Orexin/hypocretin excites the histaminergic neurons of the tuberomammillary nucleus. J Neurosci 21: 9273-9279. |
| [25] |
Murai Y, Akaike T. (2005) Orexins cause depolarization via nonselective cationic and K+ channels in isolated locus coeruleus neurons. Neurosci Res 51: 55-65. doi: 10.1016/j.neures.2004.09.005
|
| [26] |
Yu L, Zhang XY, Zhang J, et al. (2010) Orexins excite neurons of the rat cerebellar nucleus interpositus via orexin 2 receptors in vitro. Cerebellum. 9: 88-95. doi: 10.1007/s12311-009-0146-0
|
| [27] |
Scelfo B, Sacchetti B, Strata P. (2008) Learning-related long-term potentiation of inhibitory synapses in the cerebellar cortex. Proc Natl Acad Sci USA 105: 769-774. doi: 10.1073/pnas.0706342105
|
Figures(1)

Masao Ito, Naoko Nisimaru. Cerebellar Control of Defense Reactions under Orexin-mediated Neuromodulation as a Model of Cerebellohypothalamic Interaction[J]. AIMS Neuroscience, 2014, 1(1): 89-95. doi: 10.3934/Neuroscience.2014.1.89







 DownLoad:
DownLoad: