Citation: Shancy Petsel Jacob, Chikkamenahalli Lakshminarayana Lakshmikanth, Thomas M. McIntyre, Gopal Kedihitlu Marathe. Platelet-activating factor and oxidized phosphatidylcholines do not suppress endotoxin-induced pro-inflammatory signaling among human myeloid and endothelial cells[J]. AIMS Allergy and Immunology, 2017, 1(3): 108-123. doi: 10.3934/Allergy.2017.3.108

| [1] |
Zimmerman GA, Lorant DE, McIntyre TM, et al. (1993) Juxtacrine intercellular signaling: another way to do it. Am J Resp Cell Mol Biol 9: 573–577. doi: 10.1165/ajrcmb/9.6.573

|
| [2] |
Raetz CR, Whitfield C (2002) Lipopolysaccharide endotoxins. Annu Rev Biochem 71: 635–700. doi: 10.1146/annurev.biochem.71.110601.135414
|
| [3] |
Beutler B, Rietschel ET (2003) Innate immune sensing and its roots: the story of endotoxin. Nat Rev Immunol 3: 169–176. doi: 10.1038/nri1004
|
| [4] |
Medzhitov R (2001) Toll-like receptors and innate immunity. Nat Rev Immunol 1: 135–145. doi: 10.1038/35100529
|
| [5] |
Hoffmann A, Baltimore D (2006) Circuitry of nuclear factor kappa B signaling. Immunol Rev 210: 171–186. doi: 10.1111/j.0105-2896.2006.00375.x
|
| [6] | Oeckinghaus A, Ghosh S (2009) The NF-kappa B family of transcription factors and its regulation. CSH Perspect Biol 1: a000034. |
| [7] | Weyrich AS, McIntyre TM, McEver RP, et al. (1995) Monocyte tethering by P-selectin regulates monocyte chemotactic protein-1 and tumor necrosis factor-alpha secretion. Signal integration and NF-kappa B translocation. J Clin Invest 95: 2297–2303. |
| [8] |
Yost CC, Weyrich AS, Zimmerman GA (2010) The platelet-activating factor (PAF) signaling cascade in systemic inflammatory responses. Biochimie 92: 692–697. doi: 10.1016/j.biochi.2010.02.011
|
| [9] |
Brand K, Page S, Walli AK, et al. (1997) Role of nuclear factor‐kappa B in atherogenesis. Exp Physiol 82: 297–304. doi: 10.1113/expphysiol.1997.sp004025
|
| [10] |
Baker RG, Hayden MS, Ghosh S (2011) NF-kappa B, inflammation and metabolic disease. Cell Metab 13: 11–22. doi: 10.1016/j.cmet.2010.12.008
|
| [11] |
Prescott SM, Zimmerman GA, Stafforini DM, et al. (2000) Platelet-activating factor and related lipid mediators. Annu Rev Biochem 69: 419–445. doi: 10.1146/annurev.biochem.69.1.419
|
| [12] | Mozes T, Heiligers JP, Tak CJ, et al. (1991) Platelet-activating factor is one of the mediators involved in endotoxic shock in pigs. J lipid Med 4: 309–325. |
| [13] | Koltai M, Hosford D, Braquet P (1993) PAF-induced amplification of mediator release in septic shock: prevention or downregulation by PAF antagonists. J Lipid Med 6: 183–198. |
| [14] |
Shimizu T, Honda Z, Nakamura M, et al. (1992) Platelet-activating factor receptor and signal transduction. Biochem Pharmacol 44: 1001–1008. doi: 10.1016/0006-2952(92)90360-U
|
| [15] |
Marathe GK, Davies SS, Harrison KA, et al. (1999) Inflammatory platelet-activating factor-like phospholipids in oxidized low density lipoproteins are fragmented alkyl phosphatidylcholines. J Biol Chem 274: 28395–28404. doi: 10.1074/jbc.274.40.28395
|
| [16] | Marathe GK, Harrison KA, Murphy RC, et al. (2000) Bioactive phospholipid oxidation products. Free Radical Biol Med 28: 1762–1770. |
| [17] | Smiley PL, Stremler KE, Prescott SM, et al. (1991) Oxidatively fragmented phosphatidylcholines activate human neutrophils through the receptor for platelet-activating factor. J Biol Chem 266: 11104–11110. |
| [18] |
Marathe GK, Prescott SM, Zimmerman GA, et al. (2001) Oxidized LDL contains inflammatory PAF-like phospholipids. Trends Cardiovas Med 11: 139–142. doi: 10.1016/S1050-1738(01)00100-1
|
| [19] |
Leitinger N, Watson AD, Faull KF, et al. (1997) Monocyte binding to endothelial cells induced by oxidized phospholipids present in minimally oxidized low density lipoprotein is inhibited by a platelet-activating factor receptor antagonist. Adv Expt Med Biol 433: 379–382. doi: 10.1007/978-1-4899-1810-9_82
|
| [20] |
Chen R, Chen X, Salomon RG, et al. (2009) Platelet activation by low concentrations of intact oxidized LDL particles involves the PAF receptor. Arterioscl Throm Vas Biol 29: 363–371. doi: 10.1161/ATVBAHA.108.178731
|
| [21] |
Subbanagounder G, Leitinger N, Shih PT, et al. (1999) Evidence that phospholipid oxidation products and/or platelet-activating factor play an important role in early atherogenesis: in vitro and in vivo inhibition by WEB-2086. Circ Res 85: 311–318. doi: 10.1161/01.RES.85.4.311
|
| [22] |
Knapp S, Matt U, Leitinger N, et al. (2007) Oxidized phospholipids inhibit phagocytosis and impair outcome in Gram-negative sepsis in vivo. J Immunol 178: 993–1001. doi: 10.4049/jimmunol.178.2.993
|
| [23] |
Marathe GK, Pandit C, Lakshmikanth CL, et al. (2014) To hydrolyze or not to hydrolyze: the dilemma of platelet-activating factor acetylhydrolase. J Lipid Res 55: 1847–1854. doi: 10.1194/jlr.R045492
|
| [24] |
Watson AD, Leitinger N, Navab M, et al. (1997) Structural identification by mass spectrometry of oxidized phospholipids in minimally oxidized low density lipoprotein that induce monocyte/ endothelial interactions and evidence for their presence in vivo. J Biol Chem 272: 13597–13607. doi: 10.1074/jbc.272.21.13597
|
| [25] | Steinberg D, Parthasarathy S, Carew TE, et al. (1989) Beyond cholesterol. Modifications of low-density lipoprotein that increase its atherogenicity. New Engl J Med 320: 915–924. |
| [26] |
Berliner JA, Heinecke JW (1996) The role of oxidized lipoproteins in atherogenesis. Free Radical Bio Med 20: 707–727. doi: 10.1016/0891-5849(95)02173-6
|
| [27] |
Yla-Herttuala S, Palinski W, Rosenfeld ME, et al. (1989) Evidence for the presence of oxidatively modified low density lipoprotein in atherosclerotic lesions of rabbit and man. J Clin Invest 84: 1086–1095. doi: 10.1172/JCI114271
|
| [28] |
Yang L, Latchoumycandane C, McMullen MR, et al. (2010) Chronic alcohol exposure increases circulating bioactive oxidized phospholipids. J Biol Chem 285: 22211–22220. doi: 10.1074/jbc.M110.119982
|
| [29] |
Lehr HA, Weyrich AS, Saetzler RK, et al. (1997) Vitamin C blocks inflammatory platelet-activating factor mimetics created by cigarette smoking. J Clin Invest 99: 2358–2364. doi: 10.1172/JCI119417
|
| [30] |
Appleyard CB, Hillier K (1995) Biosynthesis of platelet-activating factor in normal and inflamed human colon mucosa: evidence for the involvement of the pathway of platelet- activating factor synthesis denovo in inflammatory bowel disease. Clin Sci 88: 713–717. doi: 10.1042/cs0880713
|
| [31] |
Marathe GK, Johnson C, Billings SD, et al. (2005) Ultraviolet B radiation generates platelet-activating factor-like phospholipids underlying cutaneous damage. J Biol Chem 280: 35448–35457. doi: 10.1074/jbc.M503811200
|
| [32] |
Stafforini DM (2009) Biology of platelet-activating factor acetylhydrolase (PAF-AH, lipoprotein associated phospholipase A2). Cardiovasc Drug Ther 23: 73–83. doi: 10.1007/s10557-008-6133-8
|
| [33] |
Bochkov VN, Philippova M, Oskolkova O, et al. (2006) Oxidized phospholipids stimulate angiogenesis via autocrine mechanisms, implicating a novel role for lipid oxidation in the evolution of atherosclerotic lesions. Circ Res 99: 900–908. doi: 10.1161/01.RES.0000245485.04489.ee
|
| [34] |
Silva AR, de Assis EF, Caiado LF, et al. (2002) Monocyte chemoattractant protein-1 and 5-lipoxygenase products recruit leukocytes in response to platelet-activating factor-like lipids in oxidized low-density lipoprotein. J Immunol 168: 4112–4120. doi: 10.4049/jimmunol.168.8.4112
|
| [35] |
Pontsler AV, StHilaire A, Marathe GK, et al. (2002) Cyclooxygenase-2 is induced in monocytes by peroxisome proliferator activated receptor gamma and oxidized alkyl phospholipids from oxidized low density lipoprotein. J Biol Chem 277: 13029–13036. doi: 10.1074/jbc.M109546200
|
| [36] | Ricote M, Huang J, Fajas L, et al. (1998) Expression of the peroxisome proliferator-activated receptor gamma (PPAR-gamma) in human atherosclerosis and regulation in macrophages by colony stimulating factors and oxidized low density lipoprotein. P Natl Acad Sci USA 95: 7614–7619. |
| [37] |
Schonbeck U, Sukhova GK, Graber P, et al. (1999) Augmented expression of cyclooxygenase-2 in human atherosclerotic lesions. Am J Pathol 155: 1281–1291. doi: 10.1016/S0002-9440(10)65230-3
|
| [38] |
Marathe GK, Zimmerman GA, Prescott SM, et al. (2002) Activation of vascular cells by PAF-like lipids in oxidized LDL. Vasc Pharmacol 38: 193–200. doi: 10.1016/S1537-1891(02)00169-6
|
| [39] |
Latchoumycandane C, Marathe GK, Zhang R, et al. (2012) Oxidatively truncated phospholipids are required agents of tumor necrosis factor alpha (TNF alpha)-induced apoptosis. J Biol Chem 287: 17693–17705. doi: 10.1074/jbc.M111.300012
|
| [40] |
Bochkov VN, Kadl A, Huber J, et al. (2002) Protective role of phospholipid oxidation products in endotoxin-induced tissue damage. Nature 419: 77–81. doi: 10.1038/nature01023
|
| [41] |
Jeong YI, Jung ID, Lee CM, et al. (2009) The novel role of platelet-activating factor in protecting mice against lipopolysaccharide-induced endotoxic shock. PLoS One 4: e6503. doi: 10.1371/journal.pone.0006503
|
| [42] |
Eligini S, Brambilla M, Banfi C, et al. (2002) Oxidized phospholipids inhibit cyclooxygenase-2 in human macrophages via nuclear factor-kappa B/I kappa B and ERK2-dependent mechanisms. Cardiovasc Res 55: 406–415. doi: 10.1016/S0008-6363(02)00437-6
|
| [43] |
Oskolkova OV, Afonyushkin T, Preinerstorfer B, et al. (2010) Oxidized phospholipids are more potent antagonists of lipopolysaccharide than inducers of inflammation. J Immunol 185: 7706–7712. doi: 10.4049/jimmunol.0903594
|
| [44] |
Zimmerman GA, Renzetti AD, Hill HR (1984) Granulocyte adherence in pulmonary and systemic arterial blood samples from patients with adult respiratory distress syndrome. Am Rev Respir Dis 129: 798–804. doi: 10.1164/arrd.1984.129.5.798
|
| [45] |
Smith WL, DeWitt DL, Garavito RM (2000) Cyclooxygenases: structural, cellular and molecularbiology. Annu Rev Biochem 69: 145–182. doi: 10.1146/annurev.biochem.69.1.145
|
| [46] |
Herschman HR (1996) Prostaglandin synthase 2. Biochim Biophys Acta 1299: 125–140. doi: 10.1016/0005-2760(95)00194-8
|
| [47] | Maloney CG, Kutchera WA, Albertine KH, et al. (1998) Inflammatory agonists induce cyclooxygenase type 2 expression by human neutrophils. J Immunol 160: 1402–1410. |
| [48] |
Marnett LJ, Rowlinson SW, Goodwin DC, et al. (1999) Arachidonic acid oxygenation by COX-1 and COX-2 Mechanisms of catalysis and inhibition. J Biol Chem 274: 22903–22906. doi: 10.1074/jbc.274.33.22903
|
| [49] | Hoffmann E, Dittrich-Breiholz O, Holtmann H, et al. (2002) Multiple control of interleukin-8 gene expression. J Leukocyte Biol 72: 847–855. |
| [50] |
Countryman NB, Pei Y, Yi Q, et al. (2000) Evidence for involvement of the epidermal platelet-activating factor receptor in ultraviolet-B-radiation-induced interleukin-8 production. J Invest Dermatol 115: 267–272. doi: 10.1046/j.1523-1747.2000.00058.x
|
| [51] |
Yost CC, Denis MM, Lindemann S, et al. (2004) Activated polymorphonuclear leukocytes rapidly synthesize retinoic acid receptor-alpha: a mechanism for translational control of transcriptional events. J Exp Med 200: 671–680. doi: 10.1084/jem.20040224
|
| [52] |
Matt U, Sharif O, Martins R, et al. (2015) Accumulating evidence for a role of oxidized phospholipids in infectious diseases. Cell Mol Life Sci 72: 1059–1071. doi: 10.1007/s00018-014-1780-3
|
| [53] |
Mathiak G, Szewczyk D, Abdullah F, et al. (1997) Platelet-activating factor (PAF) in experimental and clinical sepsis. Shock 7: 391–404. doi: 10.1097/00024382-199706000-00001
|
| [54] |
Fukuda Y, Kawashima H, Saito K, et al. (2000) Effect of human plasma-type platelet-activating factor acetylhydrolase in two anaphylactic shock models. Eur J Pharmacol 390: 203–207. doi: 10.1016/S0014-2999(99)00920-6
|
| [55] |
Giral M, Balsa D, Ferrando R, et al. (1996) Effects of UR-12633, a new antagonist of platelet-activating factor, in rodent models of endotoxic shock. Brit J Pharmacol 118: 1223–1231. doi: 10.1111/j.1476-5381.1996.tb15527.x
|
| [56] |
Suckling KE, Macphee CH (2002) Lipoprotein-associated phospholipase A2: a target directed at the atherosclerotic plaque. Expert Opin Ther Tar 6: 309–314. doi: 10.1517/14728222.6.3.309
|
| [57] |
Mullard A (2014) GSK's darapladib failures dim hopes for anti-inflammatory heart drugs. Nat Rev Drug Discov 13: 481–482. doi: 10.1038/nrd4381
|
| [58] |
Riedemann NC, Ward PA (2002) Oxidized lipid protects against sepsis. Nat Med 8: 1084–1085. doi: 10.1038/nm1002-1084
|
| [59] |
Matt U, Sharif O, Martins R, et al. (2013) WAVE 1 mediates suppression of phagocytosis by phospholipid-derived DAMPs. J Clin Invest 123: 3014–3024. doi: 10.1172/JCI60681
|
| [60] | Jacob SP, Lakshmikanth CL, Chaithra VH, et al. (2016) Lipopolysaccharide cross-tolerance delays platelet-activating factor induced sudden death in Swiss albino mice: involvement of cyclooxygenase in cross- tolerance. PLoS One: e0153282. |
| [61] | Leitinger N, Tyner TR, Oslund L, et al. (1999) Structurally similar oxidized phospholipids differentially regulate endothelial binding of monocytes and neutrophils. P Natl Acad Sci USA 96: 12010–12015. |
| [62] |
Watanabe J, Marathe GK, Neilsen PO, et al. (2003) Endotoxins stimulate neutrophil adhesion followed by synthesis and release of platelet-activating factor in microparticles. J Biol Chem 278: 33161–33168. doi: 10.1074/jbc.M305321200
|
| [63] | Stafforini DM, Elstad MR, McIntyre TM, et al. (1990) Human macrophages secret platelet-activating factor acetylhydrolase. J Biol Chem 265: 9682–9687. |
| [64] |
Uhlson C, Harrison K, Allen CB, et al. (2002) Oxidized phospholipids derived from ozone-treated lung surfactant extract reduce macrophage and epithelial cell viability. Chem Res Toxicol 15: 896–906. doi: 10.1021/tx010183i
|
| [65] |
Dahl M, Bauer AK, Arredouani M, et al. (2007) Protection against inhaled oxidants through scavenging of oxidized lipids by macrophage receptors MARCO and SR-AI/II. J Clin Invest 117: 757–764. doi: 10.1172/JCI29968
|
| [66] |
Podrez EA, Febbraio M, Sheibani N, et al. (2000) Macrophage scavenger receptor CD36 is the major receptor for LDL modified by monocyte-generated reactive nitrogen species. J Clin Invest 105: 1095–1108. doi: 10.1172/JCI8574
|
| [67] |
Chen M, Masaki T, Sawamura T (2002) LOX-1, the receptor for oxidized low-density lipoprotein identified from endothelial cells: implications in endothelial dysfunction and atherosclerosis. Pharmacol Therapeut 95: 89–100. doi: 10.1016/S0163-7258(02)00236-X
|
| [68] |
Teixeira-da-Cunha MG, Gomes RN, Roehrs N, et al. (2013) Bacterial clearance is improved in septic mice by platelet-activating factor-acetylhydrolase (PAF-AH) administration. PLoS One 8: e74567. doi: 10.1371/journal.pone.0074567
|
| [69] |
Chen CH, Jiang T, Yang JH, et al. (2003) Low-density lipoprotein in hypercholesterolemic human plasma induces vascular endothelial cell apoptosis by inhibiting fibroblast growth factor 2 transcription. Circulation 107: 2102–2108. doi: 10.1161/01.CIR.0000065220.70220.F7
|
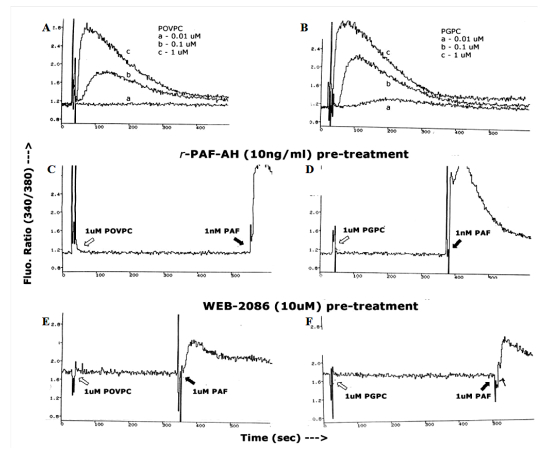
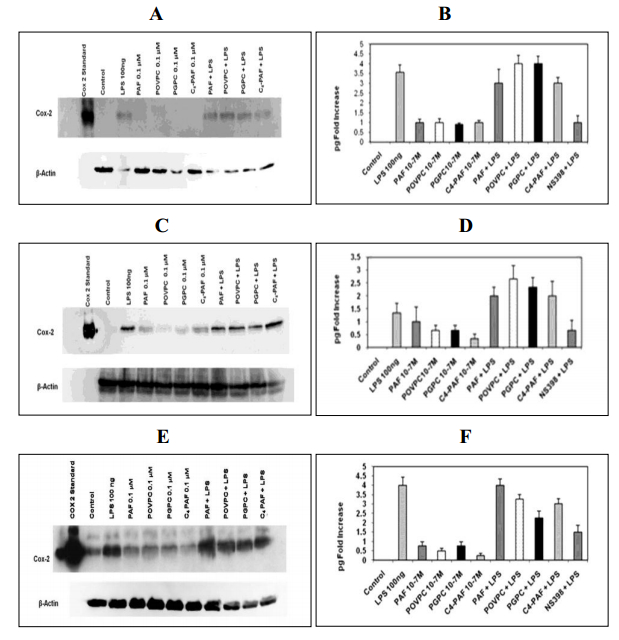
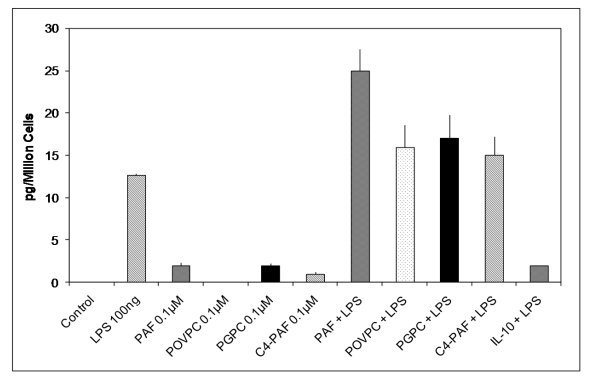
Figures(3)

Shancy Petsel Jacob, Chikkamenahalli Lakshminarayana Lakshmikanth, Thomas M. McIntyre, Gopal Kedihitlu Marathe. Platelet-activating factor and oxidized phosphatidylcholines do not suppress endotoxin-induced pro-inflammatory signaling among human myeloid and endothelial cells[J]. AIMS Allergy and Immunology, 2017, 1(3): 108-123. doi: 10.3934/Allergy.2017.3.108







 DownLoad:
DownLoad: