Citation: Ajay Pradhan, Per-Erik Olsson. Regulation of zebrafish gonadal sex differentiation[J]. AIMS Molecular Science, 2016, 3(4): 567-584. doi: 10.3934/molsci.2016.4.567

| [1] |
Dooley K, Zon LI (2000) Zebrafish: a model system for the study of human disease. Curr Opin Genet Dev 10: 252-256. doi: 10.1016/S0959-437X(00)00074-5

|
| [2] | Goldsmith JR, Jobin C (2012) Think Small: Zebrafish as a Model System of Human Pathology. J Biomed Biotechnol 2012: 817341. |
| [3] |
Lieschke GJ, Currie PD (2007) Animal models of human disease: zebrafish swim into view. Nat Rev Genet 8: 353-367. doi: 10.1038/nrg2091
|
| [4] |
Mandrekar N, Thakur NL (2009) Significance of the zebrafish model in the discovery of bioactive molecules from nature. Biotechnol Lett 31: 171-179. doi: 10.1007/s10529-008-9868-1
|
| [5] |
Zon LI, Peterson RT (2005) In vivo drug discovery in the zebrafish. Nat Rev Drug Discov 4: 35-44. doi: 10.1038/nrd1606
|
| [6] |
Skakkebaek NE, Jorgensen N, Main KM, et al. (2006) Is human fecundity declining? Int J Androl 29: 2-11. doi: 10.1111/j.1365-2605.2005.00573.x
|
| [7] |
Dumesic DA, Abbott DH, Padmanabhan V (2007) Polycystic ovary syndrome and its developmental origins. Rev Endocr Metab Disord 8: 127-141. doi: 10.1007/s11154-007-9046-0
|
| [8] | van der Zwan YG, Biermann K, Wolffenbuttel KP, et al. (2014) Gonadal Maldevelopment as Risk Factor for Germ Cell Cancer: Towards a Clinical Decision Model. Eur Urol 67: 692-701. |
| [9] |
Wilhelm D, Palmer S, Koopman P (2007) Sex determination and gonadal development in mammals. Physiol Rev 87: 1-28. doi: 10.1152/physrev.00009.2006
|
| [10] |
Angelopoulou R, Lavranos G, Manolakou P (2012) Sex determination strategies in 2012: towards a common regulatory model? Reprod Biol Endocrinol 10: 13. doi: 10.1186/1477-7827-10-13
|
| [11] | Devlin RH, Nagahama Y (2002) Sex determination and sex differentiation in fish: an overview of genetic, physiological, and environmental influences. Aquaculture 208: 191-364. |
| [12] |
Liew WC, Bartfai R, Lim Z, et al. (2012) Polygenic sex determination system in zebrafish. PLoS One 7: e34397. doi: 10.1371/journal.pone.0034397
|
| [13] | Liew WC, Orban L (2013) Zebrafish sex: a complicated affair. Brief Funct Genom 13: 172-187. |
| [14] |
Ser JR, Roberts RB, Kocher TD (2010) Multiple interacting loci control sex determination in lake Malawi cichlid fish. Evolution 64: 486-501. doi: 10.1111/j.1558-5646.2009.00871.x
|
| [15] | Vandeputte M, Dupont-Nivet M, Chavanne H, et al. (2007) A polygenic hypothesis for sex determination in the European sea bass Dicentrarchus labrax. Genetics 176: 1049-1057. |
| [16] |
Matsuda M, Nagahama Y, Shinomiya A, et al. (2002) DMY is a Y-specific DM-domain gene required for male development in the medaka fish. Nature 417: 559-563. doi: 10.1038/nature751
|
| [17] |
Barske LA, Capel B (2008) Blurring the edges in vertebrate sex determination. Curr Opin Genet Dev 18: 499-505. doi: 10.1016/j.gde.2008.11.004
|
| [18] |
Sato E, Endo T, Yamahira K, et al. (2005) Induction of female-to-male sex reversal by high temperature treatment in Medaka, Oryzias latipes. Zoolog Sci 22: 985-988. doi: 10.2108/zsj.22.985
|
| [19] |
Ospina-Alvarez N, Piferrer F (2008) Temperature-dependent sex determination in fish revisited: prevalence, a single sex ratio response pattern, and possible effects of climate change. PLoS One 3: e2837. doi: 10.1371/journal.pone.0002837
|
| [20] | Takahashi H (1977) Juvenile Hermaphroditism in the Zebrafish, Brachydanio rerio. Bull Fac Fish Hokkaido Univ 28: 57-65. |
| [21] |
Wang XG, Bartfai R, Sleptsova-Freidrich I, et al. (2007) The timing and extent of 'juvenile ovary' phase are highly variable during zebrafish testis differentiation. J Fish Biol 70: 33-44. doi: 10.1111/j.1095-8649.2007.01363.x
|
| [22] |
Siegfried KR, Nusslein-Volhard C (2008) Germ line control of female sex determination in zebrafish. Dev Biol 324: 277-287. doi: 10.1016/j.ydbio.2008.09.025
|
| [23] |
Luzio A, Monteiro SM, Garcia-Santos S, et al. (2015) Zebrafish sex differentiation and gonad development after exposure to 17alpha-ethinylestradiol, fadrozole and their binary mixture: A stereological study. Aquat Toxicol 166: 83-95. doi: 10.1016/j.aquatox.2015.07.015
|
| [24] | Uchida D, Yamashita M, Kitano T, et al. (2002) Oocyte apoptosis during the transition from ovary-like tissue to testes during sex differentiation of juvenile zebrafish. J Exp Biol 205: 711-718. |
| [25] |
Pradhan A, Khalaf H, Ochsner SA, et al. (2012) Activation of NF-kappaB protein prevents the transition from juvenile ovary to testis and promotes ovarian development in zebrafish. J Biol Chem 287: 37926-37938. doi: 10.1074/jbc.M112.386284
|
| [26] |
Sola L, Gornung E (2001) Classical and molecular cytogenetics of the zebrafish, Danio rerio (Cyprinidae, Cypriniformes): an overview. Genetica 111: 397-412. doi: 10.1023/A:1013776323077
|
| [27] |
Wallace B-M, Wallace H (2003) Synaptonemal complex karyotype of zebrafish. Heredity 90: 136-140. doi: 10.1038/sj.hdy.6800184
|
| [28] |
Traut W, Winking H (2001) Meiotic chromosomes and stages of sex chromosome evolution in fish: zebrafish, platypus and guppy. Chromosome Res 9: 659-672. doi: 10.1023/A:1012956324417
|
| [29] |
Phillips RB, Reed KM (2000) Localization of repetitive DNAs to zebrafish (Danio rerio) chromosomes by fluorescence in situ hybridization (FISH). Chromosome Res 8: 27-35. doi: 10.1023/A:1009271017998
|
| [30] | Singer A, Perlman H, Yan Y, et al. (2002) Sex-specific recombination rates in zebrafish (Danio rerio). Genetics 160: 649-657. |
| [31] |
Tong SK, Hsu HJ, Chung BC (2010) Zebrafish monosex population reveals female dominance in sex determination and earliest events of gonad differentiation. Dev Biol 344: 849-856. doi: 10.1016/j.ydbio.2010.05.515
|
| [32] | Sharma KK, Sharma OP, Tripathi NK (1998) Female heterogamety in Danio rerio (Cypriniformes: Cyprinidae). Proc Natl Acad Sci India Sect B 68: 123-126. |
| [33] |
Wilson CA, High SK, McCluskey BM, et al. (2014) Wild sex in zebrafish: loss of the natural sex determinant in domesticated strains. Genetics 198: 1291-1308. doi: 10.1534/genetics.114.169284
|
| [34] |
Shang EH, Yu RM, Wu RS (2006) Hypoxia affects sex differentiation and development, leading to a male-dominated population in zebrafish (Danio rerio). Environ Sci Technol 40: 3118-3122. doi: 10.1021/es0522579
|
| [35] |
Villamizar N, Ribas L, Piferrer F, et al. (2012) Impact of daily thermocycles on hatching rhythms, larval performance and sex differentiation of zebrafish. PLoS One 7: e52153. doi: 10.1371/journal.pone.0052153
|
| [36] |
Luzio A, Santos D, Fontainhas-Fernandes AA, et al. (2016) Effects of 17alpha-ethinylestradiol at different water temperatures on zebrafish sex differentiation and gonad development. Aquat Toxicol 174: 22-35. doi: 10.1016/j.aquatox.2016.02.003
|
| [37] |
Bradley KM, Breyer JP, Melville DB, et al. (2011) An SNP-based linkage map for zebrafish reveals sex determination loci. G3 (Bethesda) 1: 3-9. doi: 10.1534/g3.111.000190
|
| [38] |
Anderson JL, Rodriguez Mari A, Braasch I, et al. (2012) Multiple sex-associated regions and a putative sex chromosome in zebrafish revealed by RAD mapping and population genomics. PLoS One 7: e40701. doi: 10.1371/journal.pone.0040701
|
| [39] |
Mishima Y, Giraldez AJ, Takeda Y, et al. (2006) Differential regulation of germline mRNAs in soma and germ cells by zebrafish miR-430. Curr Biol 16: 2135-2142. doi: 10.1016/j.cub.2006.08.086
|
| [40] |
Staton AA, Knaut H, Giraldez AJ (2011) miRNA regulation of Sdf1 chemokine signaling provides genetic robustness to germ cell migration. Nat Genet 43: 204-211. doi: 10.1038/ng.758
|
| [41] |
von Hofsten J, Olsson PE (2005) Zebrafish sex determination and differentiation: involvement of FTZ-F1 genes. Reprod Biol Endocrinol 3: 63. doi: 10.1186/1477-7827-3-63
|
| [42] |
Orban L, Sreenivasan R, Olsson PE (2009) Long and winding roads: testis differentiation in zebrafish. Mol Cell Endocrinol 312: 35-41. doi: 10.1016/j.mce.2009.04.014
|
| [43] | Rodriguez-Mari A, Yan YL, Bremiller RA, et al. (2005) Characterization and expression pattern of zebrafish Anti-Mullerian hormone (Amh) relative to sox9a, sox9b, and cyp19a1a, during gonad development. Gene Expr Patterns 5: 655-667. |
| [44] |
Luzio A, Coimbra AM, Benito C, et al. (2015) Screening and identification of potential sex-associated sequences in Danio rerio. Mol Reprod Dev 82: 756-764. doi: 10.1002/mrd.22508
|
| [45] |
Sim H, Argentaro A, Harley VR (2008) Boys, girls and shuttling of SRY and SOX9. Trends Endocrinol Metab 19: 213-222. doi: 10.1016/j.tem.2008.04.002
|
| [46] |
Biason-Lauber A (2010) Control of sex development. Best Pract Res Clin Endocrinol Metab 24: 163-186. doi: 10.1016/j.beem.2009.12.002
|
| [47] |
Koopman P (2005) Sex determination: a tale of two Sox genes. Trends Genet 21: 367-370. doi: 10.1016/j.tig.2005.05.006
|
| [48] |
Sekido R, Lovell-Badge R (2009) Sex determination and SRY: down to a wink and a nudge? Trends Genet 25: 19-29. doi: 10.1016/j.tig.2008.10.008
|
| [49] |
Ross AJ, Capel B (2005) Signaling at the crossroads of gonad development. Trends Endocrinol Metab 16: 19-25. doi: 10.1016/j.tem.2004.11.004
|
| [50] |
Jakob S, Lovell-Badge R (2011) Sex determination and the control of Sox9 expression in mammals. FEBS J 278: 1002-1009. doi: 10.1111/j.1742-4658.2011.08029.x
|
| [51] |
Kocer A, Reichmann J, Best D, et al. (2009) Germ cell sex determination in mammals. Mol Hum Reprod 15: 205-213. doi: 10.1093/molehr/gap008
|
| [52] |
Sreenivasan R, Cai M, Bartfai R, et al. (2008) Transcriptomic analyses reveal novel genes with sexually dimorphic expression in the zebrafish gonad and brain. PLoS One 3: e1791. doi: 10.1371/journal.pone.0001791
|
| [53] |
Jorgensen A, Morthorst JE, Andersen O, et al. (2008) Expression profiles for six zebrafish genes during gonadal sex differentiation. Reprod Biol Endocrinol 6: 25. doi: 10.1186/1477-7827-6-25
|
| [54] |
Bowles J, Feng CW, Spiller C, et al. (2010) FGF9 suppresses meiosis and promotes male germ cell fate in mice. Dev Cell 19: 440-449. doi: 10.1016/j.devcel.2010.08.010
|
| [55] |
Jameson SA, Lin YT, Capel B (2012) Testis development requires the repression of Wnt4 by Fgf signaling. Dev Biol 370: 24-32. doi: 10.1016/j.ydbio.2012.06.009
|
| [56] |
Colvin JS, Green RP, Schmahl J, et al. (2001) Male-to-female sex reversal in mice lacking fibroblast growth factor 9. Cell 104: 875-889. doi: 10.1016/S0092-8674(01)00284-7
|
| [57] |
Lasala C, Carre-Eusebe D, Picard JY, et al. (2004) Subcellular and molecular mechanisms regulating anti-Mullerian hormone gene expression in mammalian and nonmammalian species. DNA Cell Biol 23: 572-585. doi: 10.1089/dna.2004.23.572
|
| [58] |
Lourenco D, Brauner R, Rybczynska M, et al. (2011) Loss-of-function mutation in GATA4 causes anomalies of human testicular development. Proc Natl Acad Sci U S A 108: 1597-1602. doi: 10.1073/pnas.1010257108
|
| [59] |
Miyamoto Y, Taniguchi H, Hamel F, et al. (2008) A GATA4/WT1 cooperation regulates transcription of genes required for mammalian sex determination and differentiation. BMC Mol Biol 9: 44. doi: 10.1186/1471-2199-9-44
|
| [60] |
Hattori RS, Murai Y, Oura M, et al. (2012) A Y-linked anti-Mullerian hormone duplication takes over a critical role in sex determination. Proc Natl Acad Sci U S A 109: 2955-2959. doi: 10.1073/pnas.1018392109
|
| [61] |
Wang XG, Orban L (2007) Anti-Mullerian hormone and 11 beta-hydroxylase show reciprocal expression to that of aromatase in the transforming gonad of zebrafish males. Dev Dyn 236: 1329-1338. doi: 10.1002/dvdy.21129
|
| [62] |
Kluver N, Pfennig F, Pala I, et al. (2007) Differential expression of anti-Mullerian hormone (amh) and anti-Mullerian hormone receptor type II (amhrII) in the teleost medaka. Dev Dyn 236: 271-281. doi: 10.1002/dvdy.20997
|
| [63] | Kamiya T, Kai W, Tasumi S, et al. (2012) A Trans-Species Missense SNP in Amhr2 Is Associated with Sex Determination in the Tiger Pufferfish, Takifugu rubripes (Fugu). PLoS Genet 8: e1002798. |
| [64] |
Pradhan A, Olsson PE (2014) Juvenile Ovary to Testis Transition in Zebrafish Involves Inhibition of Ptges. Biol Reprod 91: 33. doi: 10.1095/biolreprod.114.119016
|
| [65] | Sreenivasan R, Jiang J, Wang X, et al. (2013) Gonad Differentiation in Zebrafish Is Regulated by the Canonical Wnt Signaling Pathway. Biol Reprod 90: 45. |
| [66] |
Kazanskaya O, Glinka A, del Barco Barrantes I, et al. (2004) R-Spondin2 is a secreted activator of Wnt/beta-catenin signaling and is required for Xenopus myogenesis. Dev Cell 7: 525-534. doi: 10.1016/j.devcel.2004.07.019
|
| [67] |
Jin YR, Yoon JK (2012) The R-spondin family of proteins: emerging regulators of WNT signaling. Int J Biochem Cell Biol 44: 2278-2287. doi: 10.1016/j.biocel.2012.09.006
|
| [68] |
Zhang Y, Li F, Sun D, et al. (2011) Molecular analysis shows differential expression of R-spondin1 in zebrafish (Danio rerio) gonads. Mol Biol Rep 38: 275-282. doi: 10.1007/s11033-010-0105-3
|
| [69] |
Smith CA, Shoemaker CM, Roeszler KN, et al. (2008) Cloning and expression of R-Spondin1 in different vertebrates suggests a conserved role in ovarian development. BMC Dev Biol 8: 72. doi: 10.1186/1471-213X-8-72
|
| [70] |
Yoon JK, Lee JS (2012) Cellular signaling and biological functions of R-spondins. Cell Signal 24: 369-377. doi: 10.1016/j.cellsig.2011.09.023
|
| [71] |
Eisinger AL, Nadauld LD, Shelton DN, et al. (2007) Retinoic acid inhibits beta-catenin through suppression of Cox-2: a role for truncated adenomatous polyposis coli. J Biol Chem 282: 29394-29400. doi: 10.1074/jbc.M609768200
|
| [72] |
Goessling W, North TE, Loewer S, et al. (2009) Genetic interaction of PGE2 and Wnt signaling regulates developmental specification of stem cells and regeneration. Cell 136: 1136-1147. doi: 10.1016/j.cell.2009.01.015
|
| [73] |
Pannetier M, Fabre S, Batista F, et al. (2006) FOXL2 activates P450 aromatase gene transcription: towards a better characterization of the early steps of mammalian ovarian development. J Mol Endocrinol 36: 399-413. doi: 10.1677/jme.1.01947
|
| [74] |
Fleming NI, Knower KC, Lazarus KA, et al. (2010) Aromatase Is a Direct Target of FOXL2: C134W in Granulosa Cell Tumors via a Single Highly Conserved Binding Site in the Ovarian Specific Promoter. PLoS One 5: e14389. doi: 10.1371/journal.pone.0014389
|
| [75] |
Nef S, Vassalli JD (2009) Complementary pathways in mammalian female sex determination. J Biol 8: 74. doi: 10.1186/jbiol173
|
| [76] |
Guiguen Y, Fostier A, Piferrer F, et al. (2010) Ovarian aromatase and estrogens: a pivotal role for gonadal sex differentiation and sex change in fish. Gen Comp Endocrinol 165: 352-366. doi: 10.1016/j.ygcen.2009.03.002
|
| [77] |
Fenske M, Segner H (2004) Aromatase modulation alters gonadal differentiation in developing zebrafish (Danio rerio). Aquat Toxicol 67: 105-126. doi: 10.1016/j.aquatox.2003.10.008
|
| [78] | Takatsu K, Miyaoku K, Roy SR, et al. (2013) Induction of female-to-male sex change in adult zebrafish by aromatase inhibitor treatment. Sci Rep 3: 3400. |
| [79] |
Brion F, Tyler CR, Palazzi X, et al. (2004) Impacts of 17beta-estradiol, including environmentally relevant concentrations, on reproduction after exposure during embryo-larval-, juvenile- and adult-life stages in zebrafish (Danio rerio). Aquat Toxicol 68: 193-217. doi: 10.1016/j.aquatox.2004.01.022
|
| [80] |
Orn S, Holbech H, Norrgren L (2016) Sexual disruption in zebrafish (Danio rerio) exposed to mixtures of 17alpha-ethinylestradiol and 17beta-trenbolone. Environ Toxicol Pharmacol 41: 225-231. doi: 10.1016/j.etap.2015.12.010
|
| [81] | Schulz RW, Bogerd J, Male R, et al. (2007) Estrogen-induced alterations in amh and dmrt1 expression signal for disruption in male sexual development in the zebrafish. Environ Sci Technol 41: 6305-6310. |
| [82] |
Reyhanian Caspillo N, Volkova K, Hallgren S, et al. (2014) Short-term treatment of adult male zebrafish (Danio Rerio) with 17alpha-ethinyl estradiol affects the transcription of genes involved in development and male sex differentiation. Comp Biochem Physiol C Toxicol Pharmacol 164: 35-42. doi: 10.1016/j.cbpc.2014.04.003
|
| [83] |
Lor Y, Revak A, Weigand J, et al. (2015) Juvenile exposure to vinclozolin shifts sex ratios and impairs reproductive capacity of zebrafish. Reprod Toxicol 58: 111-118. doi: 10.1016/j.reprotox.2015.09.003
|
| [84] |
Baumann L, Knorr S, Keiter S, et al. (2014) Persistence of endocrine disruption in zebrafish (Danio rerio) after discontinued exposure to the androgen 17beta-trenbolone. Environ Toxicol Chem 33: 2488-2496. doi: 10.1002/etc.2698
|
| [85] |
Mukhi S, Torres L, Patino R (2007) Effects of larval-juvenile treatment with perchlorate and co-treatment with thyroxine on zebrafish sex ratios. Gen Comp Endocrinol 150: 486-494. doi: 10.1016/j.ygcen.2006.11.013
|
| [86] |
Sharma P, Tang S, Mayer GD, et al. (2016) Effects of thyroid endocrine manipulation on sex-related gene expression and population sex ratios in Zebrafish. Gen Comp Endocrinol 235: 38-47. doi: 10.1016/j.ygcen.2016.05.028
|
| [87] |
Sharma P, Patino R (2013) Regulation of gonadal sex ratios and pubertal development by the thyroid endocrine system in zebrafish (Danio rerio). Gen Comp Endocrinol 184: 111-119. doi: 10.1016/j.ygcen.2012.12.018
|
| [88] |
Aggarwal BB, Sethi G, Nair A, et al. (2006) Nuclear factor- κB: A holy grail in cancer prevention and therapy. Curr Signal Transduct Ther 1: 25-52. doi: 10.2174/157436206775269235
|
| [89] |
Li X, Stark GR (2002) NF-κB-dependent signaling pathways. Exp Hematol 30: 285-296. doi: 10.1016/S0301-472X(02)00777-4
|
| [90] |
Ghosh S, May MM, Kopp EB (1998) NF-κB and Rel proteins: evolutionarily conserved mediators of immune responses. Annu Rev Immunol 16: 225-260. doi: 10.1146/annurev.immunol.16.1.225
|
| [91] | Siebenlist U, Franzoso G, Brown K (1994) Structure, regulation and function of NF-κB. Annu Rev Cell Biol 10: 405-455. |
| [92] | Xiao W (2004) Advances in NF-kappaB signaling transduction and transcription. Cell Mol Immunol 1: 425-435. |
| [93] |
Beg AA, Baldwin AS (1993) The I kappa B proteins: multifunctional regulators of Rel/NF-kappa B transcription factors. Genes Dev 7: 2064-2070. doi: 10.1101/gad.7.11.2064
|
| [94] | Ghosh S, Karin M (2002) Missing pieces in the NFkB puzzle. Cell Metab 109: 81-96. |
| [95] |
Beg AA, Finco TS, Nantermet PV, et al. (1993) Tumor necrosis factor and interleukin-1 lead to phosphorylation and loss of I kappa B alpha: a mechanism for NF-kappa B activation. Mol Cell Biol 13: 3301-3310. doi: 10.1128/MCB.13.6.3301
|
| [96] |
Finco TS, Beg AA, Baldwin AS Jr (1994) Inducible phosphorylation of I kappa B alpha is not sufficient for its dissociation from NF-kappa B and is inhibited by protease inhibitors. Proc Natl Acad Sci U S A 91: 11884-11888. doi: 10.1073/pnas.91.25.11884
|
| [97] |
Shishodia S, Aggarwal BB (2002) Nuclear factor-κB activation: A question of life or death. J Biochem Mol Biol 35: 28-40. doi: 10.5483/BMBRep.2002.35.1.028
|
| [98] |
Rao NA, McCalman MT, Moulos P, et al. (2011) Coactivation of GR and NFKB alters the repertoire of their binding sites and target genes. Genome Res 21: 1404-1416. doi: 10.1101/gr.118042.110
|
| [99] | Palvimo JJ, Reinikainen P, Ikonen T, et al. (1996) Mutual transcriptional interference between RelA and androgen receptor. J Biol Chem 271: 24151–24156. |
| [100] |
McKay LI, Cidlowski JA (1998) Cross-talk between Nuclear factor-kB and the steroid hormone receptors: Mechanisms of mutual antagonism. Mol Endocrinol 12: 45-56. doi: 10.1210/mend.12.1.0044
|
| [101] | Delfino F, Walker WH (1998) Stage-specific nuclear expression of NF-κB in mammalian testis. Mol Endocrinol 12: 1696-1707. |
| [102] |
Hong CY, Park JH, Seo KH, et al. (2003) Expression of MIS in the Testis Is Downregulated by Tumor Necrosis Factor Alpha through the Negative Regulation of SF-1 Transactivation by NF-κB. Mol Cell Biol 23: 6000-6012. doi: 10.1128/MCB.23.17.6000-6012.2003
|
| [103] | Sobolewski C, Cerella C, Dicato M, et al. (2010) The role of cyclooxygenase-2 in cell proliferation and cell death in human malignancies. Int J Cell Biol 2010: 215158. |
| [104] |
Simmons DL, Botting RM, Hla T (2004) Cyclooxygenase isozymes: the biology of prostaglandin synthesis and inhibition. Pharmacol Rev 56: 387-437. doi: 10.1124/pr.56.3.3
|
| [105] |
Kang YJ, Mbonye UR, DeLong CJ, et al. (2007) Regulation of intracellular cyclooxygenase levels by gene transcription and protein degradation. Prog Lipid Res 46: 108-125. doi: 10.1016/j.plipres.2007.01.001
|
| [106] |
Morita I (2002) Distinct functions of COX-1 and COX-2. Prostaglandins Other Lipid Mediat 68-69: 165-175. doi: 10.1016/S0090-6980(02)00029-1
|
| [107] | Tanabe T, Tohnai N (2002) Cyclooxygenase isozymes and their gene structures and expression. 68-69: 95-114. |
| [108] |
Klein T, Shephard P, Kleinert H, et al. (2007) Regulation of cyclooxygenase-2 expression by cyclic AMP. Biochim Biophys Acta 1773: 1605-1618. doi: 10.1016/j.bbamcr.2007.09.001
|
| [109] |
Schmedtje JF, Ji YS, Liu RN, et al. (1997) Hypoxia induces cyclooxygenase-2 via the NF-kB p65 transcription factor in human vascular endothelial cells. J Biol Chem 272: 601-608. doi: 10.1074/jbc.272.1.601
|
| [110] |
Tsatsanis C, Androulidaki A, Venihaki M, et al. (2006) Signalling networks regulating cyclooxygenase-2. Int J Biochem Cell Biol 38: 1654-1661. doi: 10.1016/j.biocel.2006.03.021
|
| [111] |
Poligone B, Baldwin AS (2001) Positive and negative regulation of NF-kappaB by COX-2: roles of different prostaglandins. J Biol Chem 276: 38658-38664. doi: 10.1074/jbc.M106599200
|
| [112] |
Grosser T, Yusuff S, Cheskis E, et al. (2002) Developmental expression of functional cyclooxygenases in zebrafish. Proc Natl Acad Sci U S A 99: 8418-8423. doi: 10.1073/pnas.112217799
|
| [113] | Adams IR, McLaren A (2002) Sexually dimorphic devlopment of mouse primordial germ cells: switch from oogenesis to spermatogenesis. Development 129: 1155-1164. |
| [114] |
Wilhelm D, Hiramatsu R, Mizusaki H, et al. (2007) SOX9 regulates prostaglandin D synthase gene transcription in vivo to ensure testis development. J Biol Chem 282: 10553-10560. doi: 10.1074/jbc.M609578200
|
| [115] |
Breyer RM, Bagdassarian CK, Myers SA, et al. (2001) Prostanoid receptors: subtypes and signaling. Annu Rev Pharmacol Toxicol 41: 661-690. doi: 10.1146/annurev.pharmtox.41.1.661
|
| [116] |
Cai Z, Kwintkiewicz J, Young ME, et al. (2007) Prostaglandin E2 increases Cyp19 expression in Rat Granulosa Cells: Implication of GATA-4. Mol Cell Endocrinol 263: 181-189. doi: 10.1016/j.mce.2006.09.012
|
| [117] |
Guan Y, Zhang Y, Schneider A, et al. (2001) Urogenital distribution of a mouse membrane-associated prostaglandin E2 synthase. Am J Physiol Renal Physiol 281: 1173-1177. doi: 10.1152/ajprenal.0116.2001
|
| [118] |
Sun T, Deng WB, Diao HL, et al. (2006) Differential expression and regulation of prostaglandin E synthases in the mouse ovary during sexual maturation and luteal development. J Endocrinol 189: 89-101. doi: 10.1677/joe.1.06147
|
| [119] |
Bayne RA, Eddie SL, Collins CS, et al. (2009) Prostaglandin E2 as a regulator of germ cells during ovarian development. J Clin Endocrinol Metab 94: 4053-4060. doi: 10.1210/jc.2009-0755
|
| [120] |
Takahashi T, Morrow JD, Wang H, et al. (2006) Cyclooxygenase-2-derived prostaglandin E(2) directs oocyte maturation by differentially influencing multiple signaling pathways. J Biol Chem 281: 37117-37129. doi: 10.1074/jbc.M608202200
|
| [121] |
Mank JE, Promislow DEL, Avise JC (2006) Evolution of alternative sex-determining mechanisms in teleost fishes. Biol J Linn Soc 87: 83-93. doi: 10.1111/j.1095-8312.2006.00558.x
|
| [122] |
Slanchev K, Stebler J, de la Cueva-Mendez G, et al. (2005) Development without germ cells: the role of the germ line in zebrafish sex differentiation. Proc Natl Acad Sci U S A 102: 4074-4079. doi: 10.1073/pnas.0407475102
|
| [123] |
Kurokawa H, Saito D, Nakamura S, et al. (2007) Germ cells are essential for sexual dimorphism in the medaka gonad. Proc Natl Acad Sci U S A 104: 16958-16963. doi: 10.1073/pnas.0609932104
|
| [124] | Tzung KW, Goto R, Saju JM, et al. (2014) Early Depletion of Primordial Germ Cells in Zebrafish Promotes Testis Formation. Stem Cell Rep 4: 61-73. |
| [125] |
McLaren A (2003) Primordial germ cells in the mouse. Dev Biol 262: 1-15. doi: 10.1016/S0012-1606(03)00214-8
|
| [126] | DiNapoli L, Capel B (2007) Germ cell depletion does not alter the morphogenesis of the fetal testis or ovary in the red-eared slider turtle (Trachemys scripta). J Exp Zool B Mol Dev Evol 308: 236-241. |
| [127] |
Goto R, Saito T, Takeda T, et al. (2012) Germ cells are not the primary factor for sexual fate determination in goldfish. Dev Biol 370: 98-109. doi: 10.1016/j.ydbio.2012.07.010
|
| [128] |
Fujimoto T, Nishimura T, Goto-Kazeto R, et al. (2010) Sexual dimorphism of gonadal structure and gene expression in germ cell-deficient loach, a teleost fish. Proc Natl Acad Sci U S A 107: 17211-17216. doi: 10.1073/pnas.1007032107
|
| [129] |
Petersen AM, Earp NC, Redmond ME, et al. (2016) Perchlorate Exposure Reduces Primordial Germ Cell Number in Female Threespine Stickleback. PLoS One 11: e0157792. doi: 10.1371/journal.pone.0157792
|
| [130] |
Dranow DB, Tucker RP, Draper BW (2013) Germ cells are required to maintain a stable sexual phenotype in adult zebrafish. Dev Biol 376: 43-50. doi: 10.1016/j.ydbio.2013.01.016
|
| [131] |
Rodriguez-Mari A, Canestro C, Bremiller RA, et al. (2010) Sex reversal in zebrafish fancl mutants is caused by Tp53-mediated germ cell apoptosis. PLoS Genet 6: e1001034. doi: 10.1371/journal.pgen.1001034
|
| [132] |
Rios-Rojas C, Bowles J, Koopman P (2015) On the role of germ cells in mammalian gonad development: quiet passengers or back-seat drivers? Reproduction 149: R181-191. doi: 10.1530/REP-14-0663
|
| [133] |
Behringer RR, Cate RL, Froelick GJ, et al. (1990) Abnormal sexual development in transgenic mice chronically expressing mullerian inhibiting substance. Nature 345: 167-170. doi: 10.1038/345167a0
|
| [134] |
Couse JF, Hewitt SC, Bunch DO, et al. (1999) Postnatal sex reversal of the ovaries in mice lacking estrogen receptors alpha and beta. Science 286: 2328-2331. doi: 10.1126/science.286.5448.2328
|
| [135] |
Maatouk DM, Mork L, Hinson A, et al. (2012) Germ cells are not required to establish the female pathway in mouse fetal gonads. PLoS One 7: e47238. doi: 10.1371/journal.pone.0047238
|
| [136] |
Molyneaux K, Wylie C (2004) Primordial germ cell migration. Int J Dev Biol 48: 537-544. doi: 10.1387/ijdb.041833km
|
| [137] | Saitou M, Yamaji M (2012) Primordial germ cells in mice. Cold Spring Harb Perspect Biol 4: 59-66. |
| [138] |
Kashimada K, Svingen T, Feng CW, et al. (2011) Antagonistic regulation of Cyp26b1 by transcription factors SOX9/SF1 and FOXL2 during gonadal development in mice. FASEB J 25: 3561-3569. doi: 10.1096/fj.11-184333
|
| [139] |
Bowles J, Knight D, Smith C, et al. (2006) Retinoid signaling determines germ cell fate in mice. Science 312: 596-600. doi: 10.1126/science.1125691
|
| [140] |
Koubova J, Menke DB, Zhou Q, et al. (2006) Retinoic acid regulates sex-specific timing of meiotic initiation in mice. Proc Natl Acad Sci U S A 103: 2474-2479. doi: 10.1073/pnas.0510813103
|
| [141] |
Moniot B, Ujjan S, Champagne J, et al. (2014) Prostaglandin D2 acts through the Dp2 receptor to influence male germ cell differentiation in the foetal mouse testis. Development 141: 3561-3571. doi: 10.1242/dev.103408
|
| [142] |
Kipp JL, Golebiowski A, Rodriguez G, et al. (2011) Gene expression profiling reveals Cyp26b1 to be an activin regulated gene involved in ovarian granulosa cell proliferation. Endocrinology 152: 303-312. doi: 10.1210/en.2010-0749
|
| [143] |
Pradhan A, Olsson PE (2015) Inhibition of retinoic acid synthesis disrupts spermatogenesis and fecundity in zebrafish. Gen Comp Endocrinol 217-218: 81-91. doi: 10.1016/j.ygcen.2015.02.002
|
| [144] |
Alsop D, Matsumoto J, Brown S, et al. (2008) Retinoid requirements in the reproduction of zebrafish. Gen Comp Endocrinol 156: 51-62. doi: 10.1016/j.ygcen.2007.11.008
|
| [145] |
Rodriguez-Mari A, Canestro C, BreMiller RA, et al. (2013) Retinoic acid metabolic genes, meiosis, and gonadal sex differentiation in zebrafish. PLoS One 8: e73951. doi: 10.1371/journal.pone.0073951
|
| [146] |
Azziz R, Marin C, Hoq L, et al. (2005) Health care-related economic burden of the polycystic ovary syndrome during the reproductive life span. J Clin Endocrinol Metab 90: 4650-4658. doi: 10.1210/jc.2005-0628
|
| [147] |
Agrawal R, Sharma S, Bekir J, et al. (2004) Prevalence of polycystic ovaries and polycystic ovary syndrome in lesbian women compared with heterosexual women. Fertil Steril 82: 1352-1357. doi: 10.1016/j.fertnstert.2004.04.041
|
| [148] |
Abbott DH, Barnett DK, Bruns CM, et al. (2005) Androgen excess fetal programming of female reproduction: a developmental aetiology for polycystic ovary syndrome? Hum Reprod Update 11: 357-374. doi: 10.1093/humupd/dmi013
|
| [149] |
Skakkebaek NE, Rajpert-De Meyts E, Main KM (2001) Testicular dysgenesis syndrome: an increasingly common developmental disorder with environmental aspects. Hum Reprod 16: 972-978. doi: 10.1093/humrep/16.5.972
|
| [150] |
Juul A, Almstrup K, Andersson AM, et al. (2014) Possible fetal determinants of male infertility. Nat Rev Endocrinol 10: 553-562. doi: 10.1038/nrendo.2014.97
|
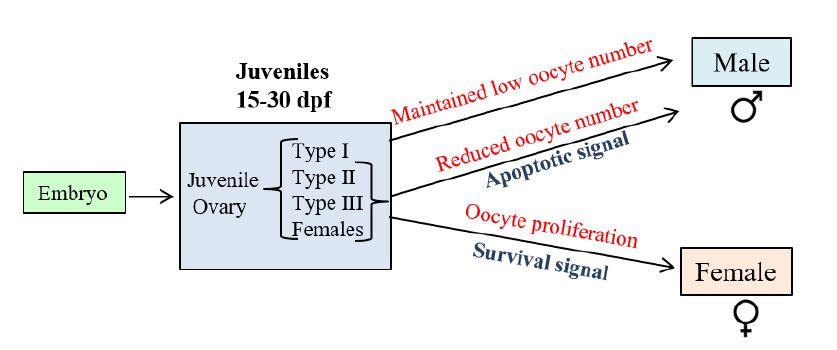
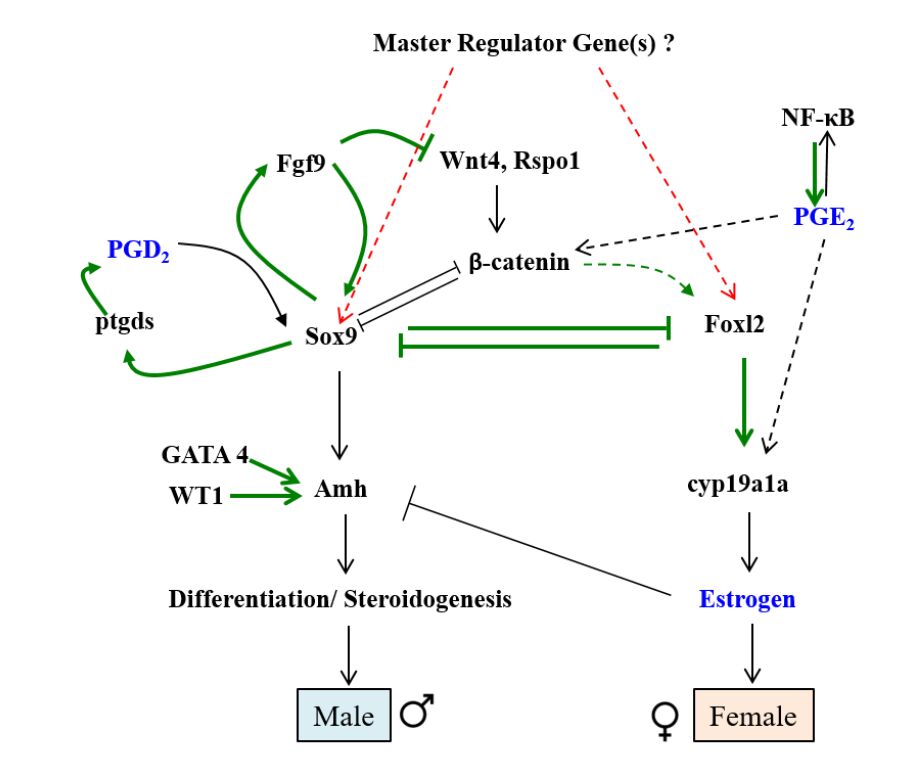
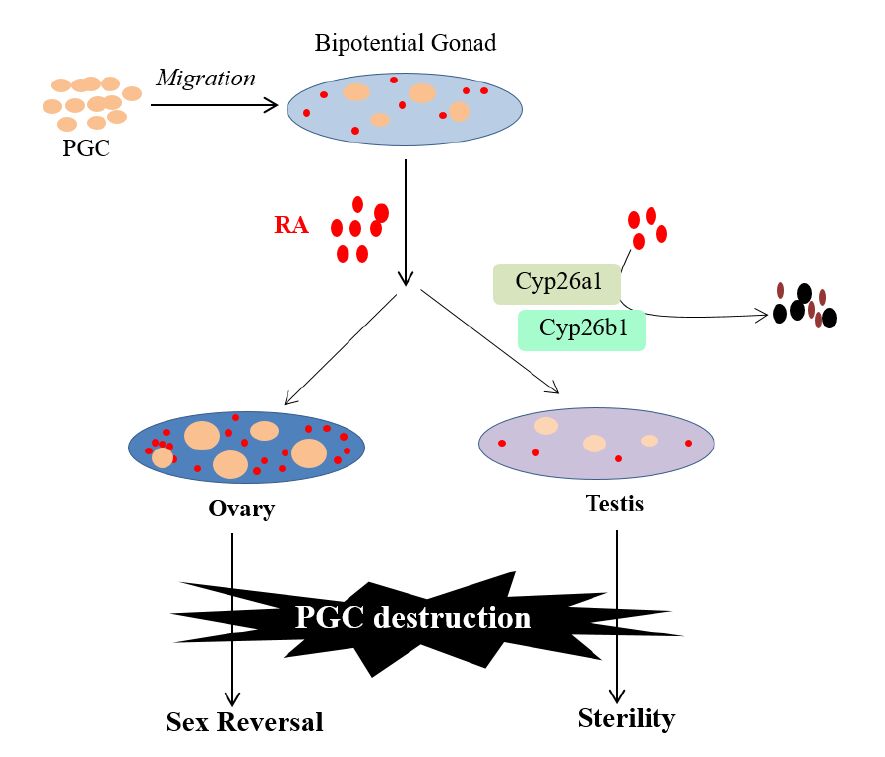
Figures(3)

Ajay Pradhan, Per-Erik Olsson. Regulation of zebrafish gonadal sex differentiation[J]. AIMS Molecular Science, 2016, 3(4): 567-584. doi: 10.3934/molsci.2016.4.567







 DownLoad:
DownLoad: