Citation: Vladimir S. Cheptsov, Elena A. Vorobyova, George A. Osipov, Natalia A. Manucharova, Lubov’ M. Polyanskaya, Mikhail V. Gorlenko, Anatoli K. Pavlov, Marina S. Rosanova, Vladimir N. Lomasov. Microbial activity in Martian analog soils after ionizing radiation: implications for the preservation of subsurface life on Mars[J]. AIMS Microbiology, 2018, 4(3): 541-562. doi: 10.3934/microbiol.2018.3.541

| [1] | Komarov IA, Isaev VS (2010) Cryology of Mars and other planets of Solar system, Moscow: Nauchnyi mir (Book in Russian). |
| [2] |
Ingersoll AP (1970) Mars: Occurrence of liquid water. Science 168: 972–973. doi: 10.1126/science.168.3934.972

|
| [3] |
Rummel JD, Beaty DW, Jones MA, et al. (2014) A new analysis of Mars "special regions": findings of the second MEPAG Special Regions Science Analysis Group (SR-SAG2). Astrobiology 14: 887–968. doi: 10.1089/ast.2014.1227
|
| [4] |
Westall F, Loizeau D, Foucher F, et al. (2013) Habitability on Mars from a microbial point of view. Astrobiology 13: 887–897. doi: 10.1089/ast.2013.1000
|
| [5] |
Ramirez RM, Craddock RA (2018) The geological and climatological case for a warmer and wetter early Mars. Nat Geosci 11: 230–237. doi: 10.1038/s41561-018-0093-9
|
| [6] |
Wordsworth R, Kalugina Y, Lokshtanov S, et al. (2017) Transient reducing greenhouse warming on early Mars. Geophys Res Lett 44: 665–671. doi: 10.1002/2016GL071766
|
| [7] |
Wordsworth RD, Kerber L, Pierrehumbert RT, et al. (2015) Comparison of "warm and wet" and "cold and icy" scenarios for early Mars in a 3-D climate model. J Geophys Res-Planet 120: 1201–1219. doi: 10.1002/2015JE004787
|
| [8] |
Nutman AP, Bennett VC, Friend CRL, et al. (2016) Rapid emergence of life shown by discovery of 3,700-million-year-old microbial structures. Nature 537: 535–538. doi: 10.1038/nature19355
|
| [9] |
Allwood AC (2016) Geology: evidence of life in Earth's oldest rocks. Nature 537: 500–501. doi: 10.1038/nature19429
|
| [10] |
Beaty DW, Clifford SM, Borg LE, et al. (2005) Key science questions from the second conference on early Mars: geologic, hydrologic, and climatic evolution and the implications for life. Astrobiology 5: 663–689. doi: 10.1089/ast.2005.5.663
|
| [11] |
Vorobyova E, Soina V, Gorlenko M, et al. (1997) The deep cold biosphere: facts and hypothesis. FEMS Microbiol Rev 20: 277–290. doi: 10.1111/j.1574-6976.1997.tb00314.x
|
| [12] |
Gilichinsky DA, Wilson GS, Friedmann EI, et al. (2007) Microbial populations in Antarctic permafrost: biodiversity, state, age and implication for astrobiology. Astrobiology 7: 275–311. doi: 10.1089/ast.2006.0012
|
| [13] | Fendrihan S, Legat A, Pfaffenhuemer M, et al. (2006) Extremely halophilic archaea and the issue of long-term microbial survival. Rev Environ Sci Bio 5: 203–218. |
| [14] |
Pavlov AK, Blinov AV, Konstantinov AN (2002) Sterilization of Martian surface by cosmic radiation. Planet Space Sci 50: 669–673. doi: 10.1016/S0032-0633(01)00113-1
|
| [15] | Halliwell B, Gutteridge JMC (2015) Free radicals in biology and medicine, New York: Oxford University Press. |
| [16] | Baumstark-Khan C, Facius R (2002) Life under conditions of ionizing radiation, In: Horneck G, Baumstark-Khan C, Editors, Astrobiology: The Quest for the Conditions of Life, Berlin: Springer, 261–284. |
| [17] | Ghosal D, Omelchenko MV, Gaidamakova EK, et al. (2005) How radiation kills cells: survival of Deinococcus radiodurans and Shewanella oneidensis under oxidative stress. FEMS Microbiol Rev 29: 361–375. |
| [18] |
Bauermeister A, Moeller R, Reitz G, et al. (2011) Effect of relative humidity on Deinococcus radiodurans' resistance to prolonged desiccation, heat, ionizing, germicidal, and environmentally relevant UV radiation. Microbial Ecol 61: 715–722. doi: 10.1007/s00248-010-9785-4
|
| [19] | Verseux C, Baqué M, Cifariello R, et al. (2017) Evaluation of the resistance of Chroococcidiopsis spp. to sparsely and densely ionizing irradiation. Astrobiology 17: 118–125. |
| [20] |
Henriksen T (1962) Radiation-induced free radicals in frozen aqueous solutions of glycine. Radiat Res 17: 158–172. doi: 10.2307/3571306
|
| [21] |
Dartnell LR, Hunter SJ, Lovell KV, et al. (2010) Low-temperature ionizing radiation resistance of Deinococcus radiodurans and Antarctic Dry Valley bacteria. Astrobiology 10: 717–732. doi: 10.1089/ast.2009.0439
|
| [22] |
Richmond R, Sridhar R, Daly MJ (1999) Physicochemical survival pattern for the radiophile Deinococcus radiodurans: a polyextremophile model for life on Mars. Proc Spie 3755: 210–222. doi: 10.1117/12.375078
|
| [23] |
Sommers CH, Niemira BA, Tunick M, et al. (2002) Effect of temperature on the radiation resistance of virulent Yersinia enterocolitica. Meat Sci 61: 323–328. doi: 10.1016/S0309-1740(01)00200-5
|
| [24] |
El-Sayed WS, Ghanem S (2009) Bacterial community community structure change induced by gamma irradiation in hydrocarbon contaminated and uncontaminated soils revealed by PCR-denaturing gradient gel electrophoresis. Biotechnology 8: 78–85. doi: 10.3923/biotech.2009.78.85
|
| [25] |
Stotzky G, Mortensen JL (1959) Effect of gamma radiation on growth and metabolism of microorganisms in an organic soil. Soil Sci Soc Am J 23: 125–127. doi: 10.2136/sssaj1959.03615995002300020014x
|
| [26] |
McNamara NP, Black HIJ, Beresford NA, et al. (2003) Effects of acute gamma irradiation on chemical, physical and biological properties of soils. Appl Soil Ecol 24: 117–132. doi: 10.1016/S0929-1393(03)00073-8
|
| [27] |
Smith HD, McKay CP (2005) Drilling in ancient permafrost on Mars for evidence of a second genesis of life. Planet Space Sci 53: 1302–1308. doi: 10.1016/j.pss.2005.07.006
|
| [28] | Wierzchos J, de los Ríos A, Ascaso C (2012) Microorganisms in desert rocks: the edge of life on Earth. Int Microbiol 15: 171–181. |
| [29] |
DiRuggiero J, Wierzchos J, Robinson CK, et al. (2013) Microbial colonisation of chasmoendolithic habitats in the hyper-arid zone of the Atacama Desert. Biogeosciences 10: 2439–2450. doi: 10.5194/bg-10-2439-2013
|
| [30] |
Parro V, de Diego-Castilla G, Moreno-Paz M, et al. (2011) A microbial oasis in the hypersaline Atacama subsurface discovered by a life detector chip: implications for the search for life on Mars. Astrobiology 11: 969–996. doi: 10.1089/ast.2011.0654
|
| [31] |
Fairén AG, Davila AF, Lim D, et al. (2010) Astrobiology through the ages of Mars: the study of terrestrial analogues to understand the habitability of Mars. Astrobiology 10: 821–843. doi: 10.1089/ast.2009.0440
|
| [32] |
Frösler J, Panitz C, Wingender J, et al. (2017) Survival of Deinococcus geothermalis in biofilms under desiccation and simulated space and Martian conditions. Astrobiology 17: 431–447. doi: 10.1089/ast.2015.1431
|
| [33] |
Decho AW (2000) Microbial biofilms in intertidal systems: an overview. Cont Shelf Res 20: 1257–1273. doi: 10.1016/S0278-4343(00)00022-4
|
| [34] | Soina VS, Vorobyova EA (2004) Adaptation of Bacteria to the Terrestrial Permafrost Environment, In: Origins, Dordrecht: Springer Netherlands, 427–444. |
| [35] |
El-Registan GI, Mulyukin AL, Nikolaev YA, et al. (2006) Adaptogenic functions of extracellular autoregulators of microorganisms. Microbiology 75: 380–389. doi: 10.1134/S0026261706040035
|
| [36] |
Sugden DE, Marchant DR, Potter N, et al. (1995) Preservation of Miocene glacier ice in East Antarctica. Nature 376: 412–414. doi: 10.1038/376412a0
|
| [37] | Schumacher BA (2002) Methods for the determination of total organic carbon (TOC) in soils and sediments, NCEA-C-1282, Washington: U.S. Environmental Protection Agency, National Exposure Research Laboratory. |
| [38] |
Groemer G, Soucek A, Frischauf N, et al. (2014) The MARS2013 Mars analog mission. Astrobiology 14: 360–376. doi: 10.1089/ast.2013.1062
|
| [39] | Impey C, Henry H (2016) Dreams of other worlds: the amazing story of unmanned space exploration, Princeton: Princeton University Press, 449. |
| [40] |
Pavlov AK, Shelegedin VN, Vdovina MA, et al. (2010) Growth of microorganisms in Martian-like shallow subsurface conditions: laboratory modeling. Int J Astrobiol 9: 51–58. doi: 10.1017/S1473550409990371
|
| [41] | Vishniac HS (1993) The microbiology of Antarctic soils, In: Friedmann EI, Editor, Antarctic microbiology, New York: Wiley-Liss, 297–341. |
| [42] |
Antony R, Krishnan, KP, Laluraj CM, et al. (2012) Diversity and physiology of culturable bacteria associated with a coastal Antarctic ice core. Microbiol Res 167: 372–380. doi: 10.1016/j.micres.2012.03.003
|
| [43] |
Manucharova NA, Vlasenko AN, Men'ko EV, et al. (2011) Specificity of the chitinolytic microbial complex of soils incubated at different temperatures. Microbiology 80: 205–215. doi: 10.1134/S002626171102010X
|
| [44] | Gorlenko MV, Kozhevin PA (1994) Differentiation of soil microbial communities by multisubstrate testing. Microbiology 63: 158–161. |
| [45] | Garland JL, Mills AL (1991) Classification and characterization of heterotrophic microbial communities on the basis of patterns of community-level sole-carbon-source utilization. Appl Environ Microbiol 57: 2351–2359. |
| [46] | Garland JL, Mills AL (1994) A community-level physiological approach for studying microbial communities, In: Ritz K, Dighton J, Giller KE, Editors, Beyond the biomass: Compositional and functional analysis of soil microbial communities, Chichester: Wiley, 77–83. |
| [47] | Gorlenko MV, Kozhevin PA (2005) Multisubstrate testing of natural microbial communities, Moscow: MAKS Press (Book in Russian). |
| [48] |
Manaeva ES, Lomovtseva NO, Kostina NV, et al. (2014) Biological activity of soils in the settlements of southern (Microtus rossiaemeridionalis) and bank (Clethrionomys glareolus) voles. Biol Bull 41: 80–88. doi: 10.1134/S1062359013040110
|
| [49] |
Shekhovtsova NV, Marakaev OA, Pervushina KA, et al. (2013) The underground organ microbial complexes of moorland spotted orchid Dactylorhiza maculata (L.) Soó (Orchidaceae). Adv Biosci Biotechnol 4: 35–42. doi: 10.4236/abb.2013.47A2005
|
| [50] |
Shekhovtsova NV, Osipov GA, Verkhovtseva NV, et al. (2003) Analysis of lipid biomarkers in rocks of the Archean crystalline basement. Proc Spie 4939: 160–168. doi: 10.1117/12.486691
|
| [51] | Turova ES, Osipov GA (1996) Structure of the microbial community transforming iron minerals contained in kaolin. Microbiology 65: 682–689. |
| [52] | Nozhevnikova A, Simankova M, Parshina S, et al. (2001) Temperature characteristics of methanogenic archaea and acetogenic bacteria isolated from cold environments. Water Sci Technol 44: 41–48. |
| [53] |
Cavicchioli R, Thomas T, Curmi PM (2000) Cold stress response in Archaea. Extremophiles 4: 321–331. doi: 10.1007/s007920070001
|
| [54] | Hoover RB, Pikuta EV (2010) Psychrophilic and psychrotolerant microbial extremophiles in polar environments, In: Bej AK, Aislabie J, Atlas RM, Editors, Polar microbiology: the ecology, biodiversity and bioremediation potential of microorganisms in extremely cold environments, Boca Raton: CRC Press, 115–156. |
| [55] |
Rivkina EM, Friedmann EI, McKay CP, et al. (2000) Metabolic activity of permafrost bacteria below the freezing point. Appl Environ Microbiol 66: 3230–3233. doi: 10.1128/AEM.66.8.3230-3233.2000
|
| [56] |
Cox MM, Battista JR (2005) Deinococcus radiodurans-the consummate survivor. Nat Rev Microbiol 3: 882–892. doi: 10.1038/nrmicro1264
|
| [57] |
Musilova M, Wright G, Ward JM, et al. (2015) Isolation of radiation-resistant bacteria from Mars analog Antarctic Dry Valleys by preselection, and the correlation between radiation and desiccation resistance. Astrobiology 15: 1076–1090. doi: 10.1089/ast.2014.1278
|
| [58] | Belbe GM, Tofana M (2010) Effects of ionizing radiation on microbiological contaminants of foods. B UASVM Agr 67: 178–185. |
| [59] |
Bruhn DF, Frank SM, Roberto FF, et al. (2009) Microbial biofilm growth on irradiated, spent nuclear fuel cladding. J Nucl Mater 384: 140–145. doi: 10.1016/j.jnucmat.2008.11.008
|
| [60] |
Zhang YM, Rock CO (2008) Membrane lipid homeostasis in bacteria. Nat Rev Microbiol 6: 222–233. doi: 10.1038/nrmicro1839
|
| [61] |
Mansilla MC, Cybulski LE, Albanesi D, et al. (2004) Control of membrane lipid fluidity by molecular thermosensors. J Bacteriol 186: 6681–6688. doi: 10.1128/JB.186.20.6681-6688.2004
|
| [62] |
Rainey FA, Ray K, Ferreira M, et al. (2005) Extensive diversity of ionizing-radiation-resistant bacteria recovered from Sonoran Desert soil and description of nine new species of the genus Deinococcus obtained from a single soil sample. Appl Environ Microbiol 71: 5225–5235. doi: 10.1128/AEM.71.9.5225-5235.2005
|
| [63] | Beblo-Vranesevic K, Koschnitzki D, Rettberg P (2016) Ignicoccus hospitalis: a polyextremophilic microorganism with unusual high radiation tolerance, In: DGDR. |
| [64] |
Hassler DM, Zeitlin C, Wimmer-Schweingruber RF, et al. (2014) Mars' surface radiation environment measured with the Mars Science Laboratory's Curiosity rover. Science 343: 1244797. doi: 10.1126/science.1244797
|
| [65] | Pavlov AA, Vasilyev G, Ostryakov VM, et al. (2012) Degradation of the organic molecules in the shallow subsurface of Mars due to irradiation by cosmic rays. Geophys Res Lett 39: L13202. |
| [66] |
Dartnell LR, Desorgher L, Ward JM, et al. (2007) Martian subsurface ionising radiation: biosignatures and geology. Biogeosciences 4: 545–558. doi: 10.5194/bg-4-545-2007
|
| [67] |
Murray JB, Muller JP, Neukum G, et al. (2005) Evidence from the Mars Express High Resolution Stereo Camera for a frozen sea close to Mars' equator. Nature 434: 352–356. doi: 10.1038/nature03379
|
| [68] |
Mustard JF, Cooper CD, Rifkin MK (2001) Evidence for recent climate change on Mars from the identification of youthful near-surface ground ice. Nature 412: 411. doi: 10.1038/35086515
|
| [69] |
Madeleine JB, Head JW, Forget F, et al. (2014) Recent ice ages on Mars: the role of radiatively active clouds and cloud microphysics. Geophys Res Lett 41: 4873–4879. doi: 10.1002/2014GL059861
|
| [70] |
Johnsson A, Reiss D, Hauber E, et al. (2014) Evidence for very recent melt-water and debris flow activity in gullies in a young mid-latitude crater on Mars. Icarus 235: 37–54. doi: 10.1016/j.icarus.2014.03.005
|
| [71] |
Dieser M, Battista JR, Christner BC (2013) DNA double-strand break repair at −15 °C. Appl Environ Microbiol 79: 7662–7668. doi: 10.1128/AEM.02845-13
|
| [72] |
Martín-Torres J, Zorzano MP, Valentin-Serrano P, et al. (2015) Transient liquid water and water activity at Gale crater on Mars. Nat Geosci 8: 357–361. doi: 10.1038/ngeo2412
|
| [73] |
Jones EG (2018) Shallow transient liquid water environments on present-day mars, and their implications for life. Acta Astronaut 146: 144–150. doi: 10.1016/j.actaastro.2018.02.027
|
| [74] |
Jones EG, Lineweaver CH, Clarke JD (2011) An extensive phase space for the potential martian biosphere. Astrobiology 11: 1017–1033. doi: 10.1089/ast.2011.0660
|
| [75] | Nicholson WL, Krivushin K, Gilichinsky D, et al. (2013) Growth of Carnobacterium spp. from permafrost under low pressure, temperature, and anoxic atmosphere has implications for Earth microbes on Mars. P Nat Acad Sci USA 110: 666–671. |
| [76] |
Nicholson WL, McCoy LE, Kerney KR, et al. (2012) Aqueous extracts of a Mars analogue regolith that mimics the Phoenix landing site do not inhibit spore germination or growth of model spacecraft contaminants Bacillus subtilis 168 and Bacillus pumilus SAFR-032. Icarus 220: 904–910. doi: 10.1016/j.icarus.2012.06.033
|
| [77] |
Beblo-Vranesevic K, Huber H, Rettberg P (2017) High tolerance of Hydrogenothermus marinus to sodium perchlorate. Front Microbiol 8: 1369. doi: 10.3389/fmicb.2017.01369
|
| [78] |
Beblo-Vranesevic K, Bohmeier M, Perras AK, et al. (2017) The responses of an anaerobic microorganism, Yersinia intermedia MASE-LG-1 to individual and combined simulated Martian stresses. PLoS One 12: e0185178. doi: 10.1371/journal.pone.0185178
|
| [79] |
Al Soudi AF, Farhat O, Chen F, et al. (2017) Bacterial growth tolerance to concentrations of chlorate and perchlorate salts relevant to Mars. Int J Astrobiol 16: 229–235. doi: 10.1017/S1473550416000434
|
| [80] |
Nuding DL, Gough RV, Venkateswaran KJ, et al. (2017) Laboratory investigations on the survival of Bacillus subtilis spores in deliquescent salt Mars analog environments. Astrobiology 17: 997–1008. doi: 10.1089/ast.2016.1545
|
| [81] | Heinz J, Schirmack J, Airo A, et al. (2018) Enhanced microbial survivability in subzero brines. Astrobiology. |
| [82] |
Koskinen K, Rettberg P, Pukall R, et al. (2017) Microbial biodiversity assessment of the European Space Agency's ExoMars 2016 mission. Microbiome 5: 143. doi: 10.1186/s40168-017-0358-3
|
| [83] | Thomas H, Rettberg P, Shimizu T, et al. (2016) Cold atmospheric plasma technology for decontamination of space equipment, In: 41st COSPAR Scientific Assembly. |
| [84] |
Hahn C, Hans M, Hein C, et al. (2017) Pure and oxidized copper materials as potential antimicrobial surfaces for spaceflight activities. Astrobiology 17: 1183–1191. doi: 10.1089/ast.2016.1620
|
| [85] |
Hecht MH, Kounaves SP, Quinn RC, et al. (2009) Detection of perchlorate and the soluble chemistry of martian soil at the Phoenix lander site. Science 325: 64–67. doi: 10.1126/science.1172466
|
| [86] |
Boynton WV, Ming DW, Kounaves SP, et al. (2009) Evidence for calcium carbonate at the Mars Phoenix landing site. Science 325: 61–64. doi: 10.1126/science.1172768
|
| [87] |
Toner JD, Catling DC, Light B (2014) Soluble salts at the Phoenix Lander site, Mars: A reanalysis of the Wet Chemistry Laboratory data. Geochim Cosmochim Ac 136: 142–168. doi: 10.1016/j.gca.2014.03.030
|
| [88] |
Meslin PY, Gasnault O, Forni O, et al. (2013) Soil diversity and hydration as observed by ChemCam at Gale Crater, Mars. Science 341: 1238670. doi: 10.1126/science.1238670
|
| [89] | Mangold N, Forni O, Dromart G, et al. (2015) Chemical variations in Yellowknife Bay formation sedimentary rocks analyzed by ChemCam on board the Curiosity rover on Mars. J Geophys Res-Planet 120: 452–482. |
| [90] |
Zakharov YA, Nevostruev VA (1968) Radiolysis of solid inorganic salts with oxygen-containing anions. Russ Chem Rev 37: 61–73. doi: 10.1070/RC1968v037n01ABEH001608
|
| [91] |
Harris DR, Pollock SV, Wood EA, et al. (2009) Directed evolution of ionizing radiation resistance in Escherichia coli. J Bacteriol 191: 5240–5252. doi: 10.1128/JB.00502-09
|
| [92] |
Tesfai AT, Beamer SK, Matak KE, et al. (2011) Radio-resistance development of DNA repair deficient Escherichia coli DH5α in ground beef subjected to electron beam at sub-lethal doses. Int J Radiat Biol 87: 571–578. doi: 10.3109/09553002.2011.565400
|
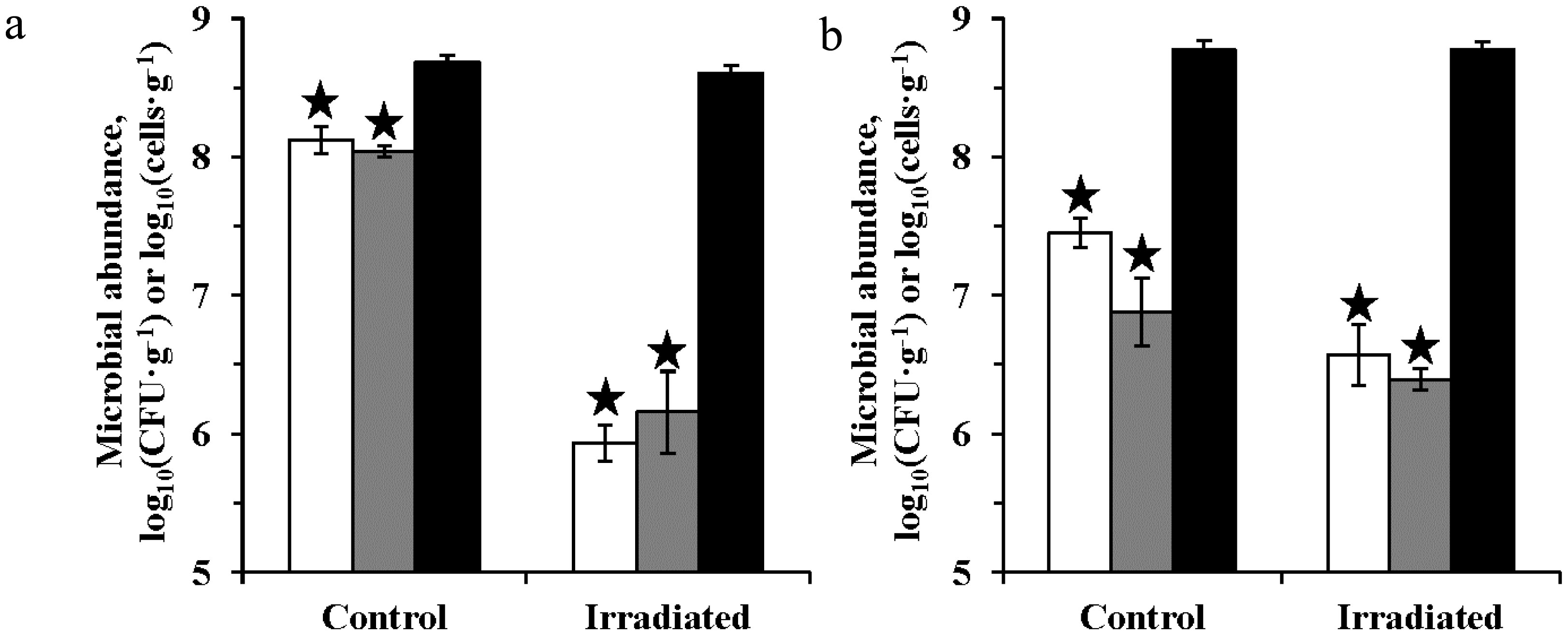
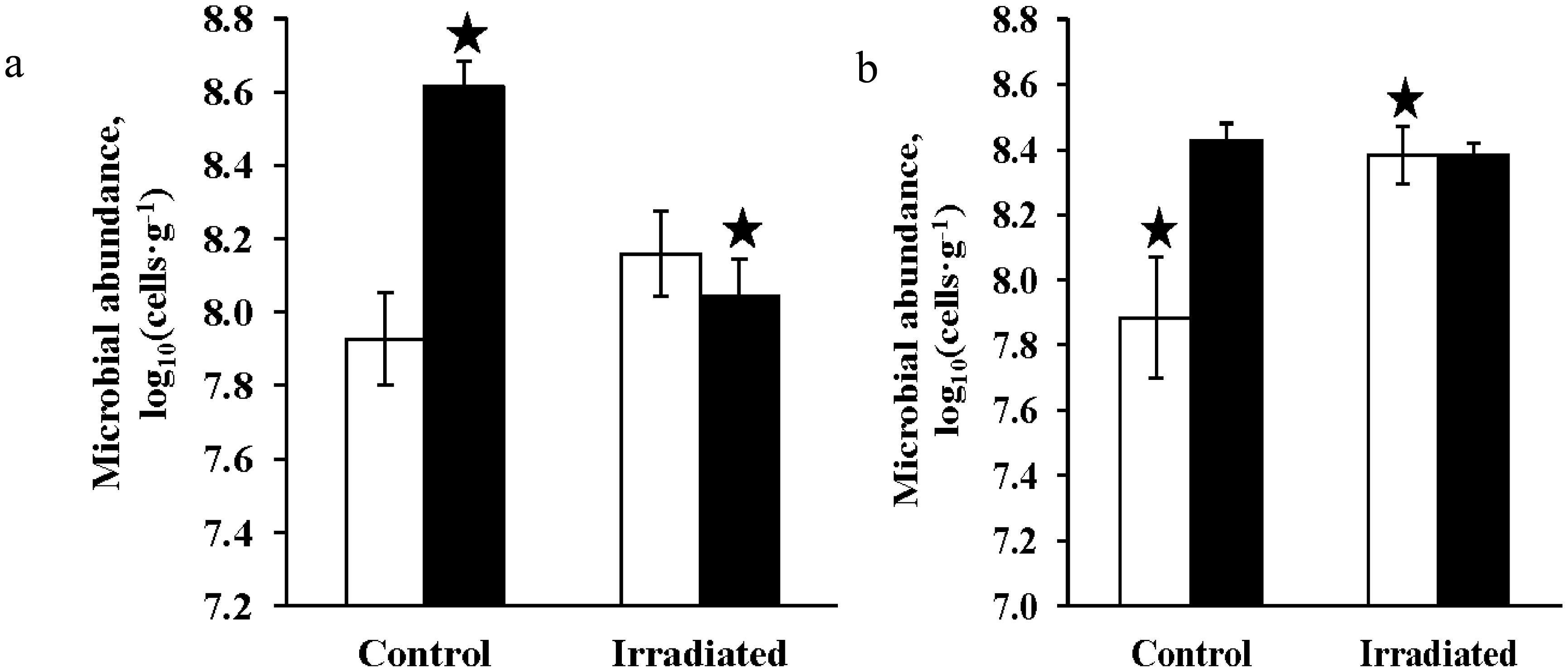
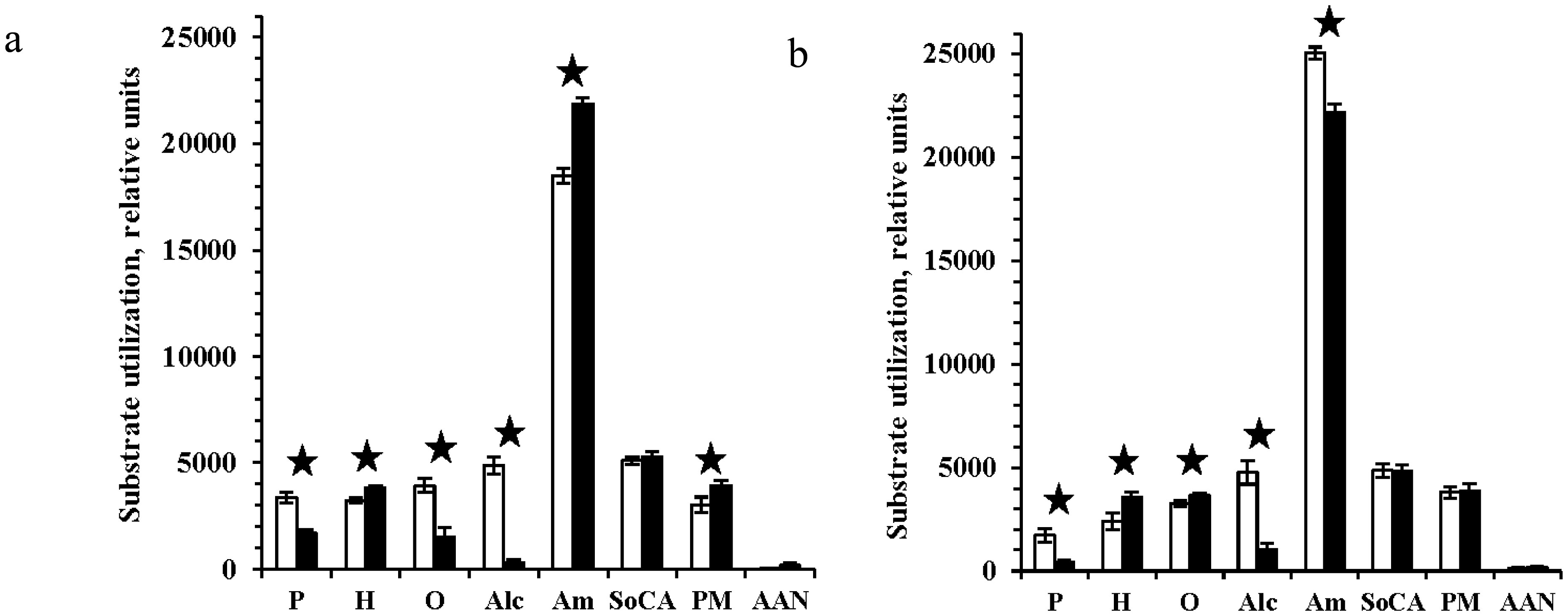
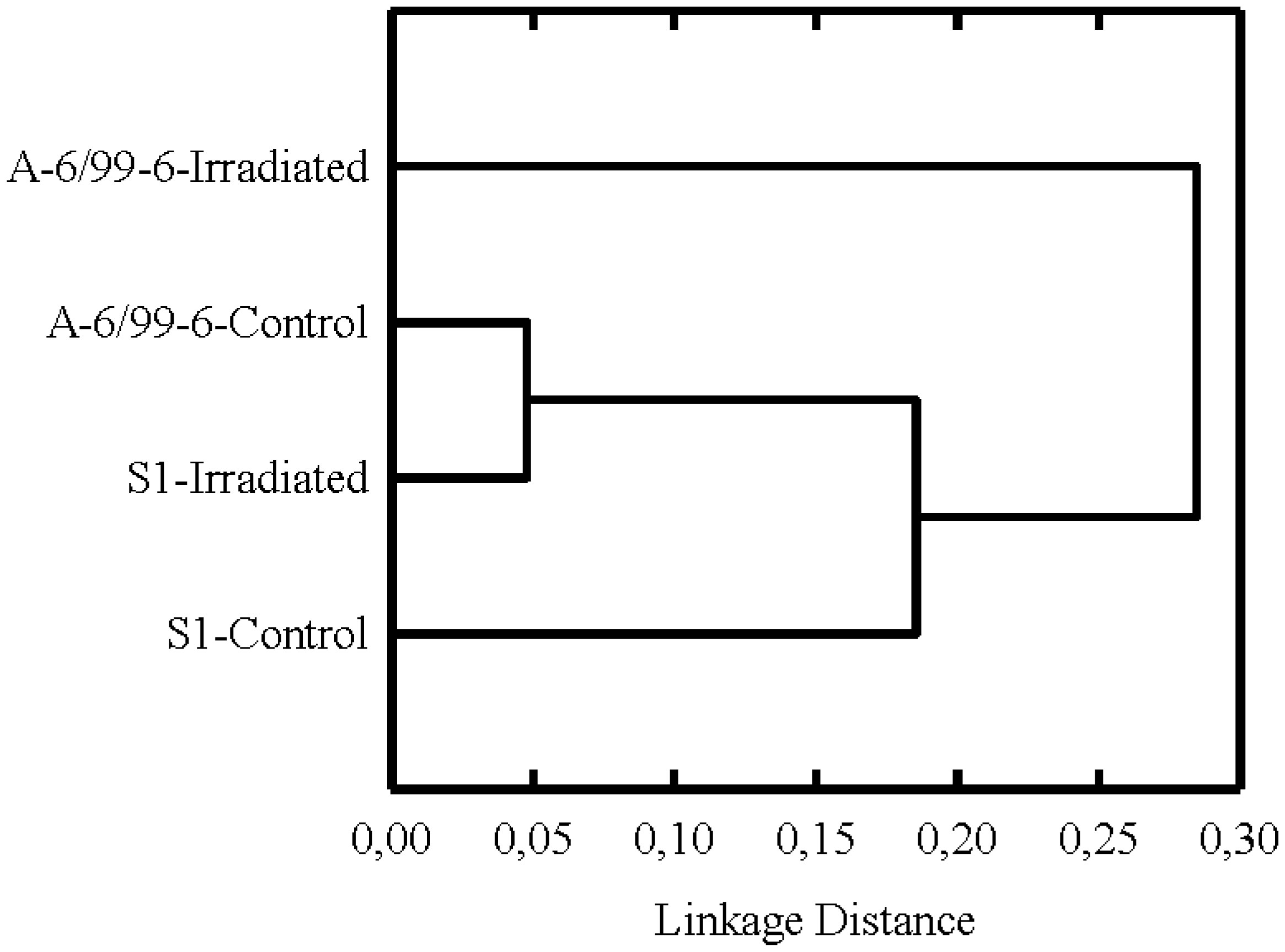
Figures(4) / Tables(6)

Vladimir S. Cheptsov, Elena A. Vorobyova, George A. Osipov, Natalia A. Manucharova, Lubov’ M. Polyanskaya, Mikhail V. Gorlenko, Anatoli K. Pavlov, Marina S. Rosanova, Vladimir N. Lomasov. Microbial activity in Martian analog soils after ionizing radiation: implications for the preservation of subsurface life on Mars[J]. AIMS Microbiology, 2018, 4(3): 541-562. doi: 10.3934/microbiol.2018.3.541







 DownLoad:
DownLoad: