Citation: Pasha Grachev, Robert L. Goodman. The GnRH Pulse Generator[J]. AIMS Medical Science, 2016, 3(4): 359-385. doi: 10.3934/medsci.2016.4.359

| [1] | Dierschke DJ, Bhattacharya AN, Atkinson LE, et al. (1970) Circhoral oscillations of plasma LH levels in the ovariectomized rhesus monkey. Endocrinology 87: 850-853. |
| [2] | Foster RG, Plowman G, Goldsmith AR, et al. (1987) Immunohistochemical demonstration of marked changes in the LHRH system of photosensitive and photorefractory European starlings (Sturnus vulgaris). J Endocrinol 115: 211-220. |
| [3] | Dalkin AC, Haisenleder DJ, Ortolano GA, et al. (1989) The frequency of gonadotropin-releasing-hormone stimulation differentially regulates gonadotropin subunit messenger ribonucleic acid expression. Endocrinology 125: 917-924. |
| [4] | Karsch FJ (1987) Central actions of ovarian steroids in the feedback regulation of pulsatile secretion of luteinizing hormone. Annu Rev Physiol 49: 365-382. |
| [5] | Herbison AE (2016) Control of puberty onset and fertility by gonadotropin-releasing hormone neurons. Nat Rev Endocrinol 12: 452-466. |
| [6] | Sarkar DK, Chiappa SA, Fink G, et al. (1976) Gonadotropin-releasing hormone surge in pro-oestrous rats. Nature 264: 461-463. |
| [7] | Moenter SM, Caraty A, Karsch FJ (1990) The estradiol-induced surge of gonadotropin-releasing hormone in the ewe. Endocrinology 127: 1375-1384. |
| [8] | Midgley AR Jr. (1966) Radioimmunoassay: a method for human chorionic gonadotropin and human luteinizing hormone. Endocrinology 79: 10-18. |
| [9] | Gay VL, Sheth NA (1972) Evidence for a periodic release of LH in castrated male and female rats. Endocrinology 90: 158-162. |
| [10] | Schally AV, Arimura A, Baba Y, et al. (1971) Isolation and properties of the FSH and LH-releasing hormone. Biochem Biophys Res Commun 43: 393-399. |
| [11] | Amoss M, Burgus R, Blackwell R, et al. (1971) Purification, amino acid composition and N-terminus of the hypothalamic luteinizing hormone releasing factor (LRF) of ovine origin. Biochem Biophys Res Commun 44: 205-210. |
| [12] | Schuiling GA, Gnodde HP (1976) Secretion of luteinizing hormone caused by continuous infusions of luteinizing hormone releasing hormone in the long-term ovariectomized rat: effect of oestrogen pretreatment. J Endocrinol 71: 1-11. |
| [13] | Belchetz PE, Plant TM, Nakai Y, et al. (1978) Hypophysial responses to continuous and intermittent delivery of hypopthalamic gonadotropin-releasing hormone. Science 202: 631-633. |
| [14] | Osland RB, Gallo RV, Williams JA (1975) In vitro release of lutenizing hormone from anterior pituitary fragments superfused with constant or pulsatile amounts of luteinizing hormone-releasing factor. Endocrinology 96: 1210-1215. |
| [15] | Nett TM, Akbar AM, Niswender GD, et al. (1973) A radioimmunoassay for gonadotropin-releasing hormone (Gn-RH) in serum. J Clin Endocrinol Metab 36: 880-885. |
| [16] | Carmel PW, Araki S, Ferin M (1976) Pituitary stalk portal blood collection in rhesus monkeys: evidence for pulsatile release of gonadotropin-releasing hormone (GnRH). Endocrinology 99: 243-248. |
| [17] | Levine JE, Ramirez VD (1980) In vivo release of luteinizing hormone-releasing hormone estimated with push-pull cannulae from the mediobasal hypothalami of ovariectomized, steroid-primed rats. Endocrinology 107: 1782-1790. |
| [18] | Clarke IJ, Cummins JT (1982) The temporal relationship between gonadotropin releasing hormone (GnRH) and luteinizing hormone (LH) secretion in ovariectomized ewes. Endocrinology 111: 1737-1739. |
| [19] | Van Vugt DA, Diefenbach WD, Alston E, et al. (1985) Gonadotropin-releasing hormone pulses in third ventricular cerebrospinal fluid of ovariectomized rhesus monkeys: correlation with luteinizing hormone pulses. Endocrinology 117: 1550-1558. |
| [20] | Halasz B, Pupp L (1965) Hormone secretion of the anterior pituitary gland after physical interruption of all nervous pathways to the hypophysiotrophic area. Endocrinology 77: 553-562. |
| [21] | Blake CA, Sawyer CH (1974) Effects of hypothalamic deafferentation on the pulsatile rhythm in plasma concentrations of luteinizing hormone in ovariectomized rats. Endocrinology 94: 730-736. |
| [22] | Krey LC, Butler WR, Knobil E (1975) Surgical disconnection of the medial basal hypothalamus and pituitary function in the rhesus monkey. I. Gonadotropin secretion. Endocrinology 96: 1073-1087. |
| [23] | Plant TM, Krey LC, Moossy J, et al. (1978) The arcuate nucleus and the control of gonadotropin and prolactin secretion in the female rhesus monkey (Macaca mulatta). Endocrinology 102: 52-62. |
| [24] | Soper BD, Weick RF (1980) Hypothalamic and extrahypothalamic mediation of pulsatile discharges of luteinizing hormone in the ovariectomized rat. Endocrinology 106: 348-355. |
| [25] | Wilson RC, Kesner JS, Kaufman JM, et al. (1984) Central electrophysiologic correlates of pulsatile luteinizing hormone secretion in the rhesus monkey. Neuroendocrinology 39: 256-260. |
| [26] | Kawakami M, Uemura T, Hayashi R (1982) Electrophysiological correlates of pulsatile gonadotropin release in rats. Neuroendocrinology 35: 63-67. |
| [27] | Thiery JC, Pelletier J (1981) Multiunit activity in the anterior median eminence and adjacent areas of the hypothalamus of the ewe in relation to LH secretion. Neuroendocrinology 32: 217-224. |
| [28] | Mori Y, Nishihara M, Tanaka T, et al. (1991) Chronic recording of electrophysiological manifestation of the hypothalamic gonadotropin-releasing hormone pulse generator activity in the goat. Neuroendocrinology 53: 392-395. |
| [29] | Silverman AJ, Antunes JL, Abrams GM, et al. (1982) The luteinizing hormone-releasing hormone pathways in rhesus (Macaca mulatta) and pigtailed (Macaca nemestrina) monkeys: new observations on thick, unembedded sections. J Comp Neurol 211: 309-317. |
| [30] | Martinez De La Escalera G, Choi AL, Weiner RI (1992) Generation and synchronization of gonadotropin-releasing hormone (GnRH) pulses: intrinsic properties of the GT1-1 GnRH neuronal cell line. Proc Natl Acad Sci U S A 89: 1852-1855. |
| [31] | Krsmanovic LZ, Stojilkovic SS, Merelli F, et al. (1992) Calcium signaling and episodic secretion of gonadotropin-releasing hormone in hypothalamic neurons. Proc Natl Acad Sci U S A 89: 8462-8466. |
| [32] | Hiruma H, Uemura T, Kimura F (1997) Neuronal synchronization and ionic mechanisms for propagation of excitation in the functional network of immortalized GT1-7 neurons: optical imaging with a voltage-sensitive dye. J Neuroendocrinol 9: 835-840. |
| [33] | Bosma MM (1993) Ion channel properties and episodic activity in isolated immortalized gonadotropin-releasing hormone (GnRH) neurons. J Membr Biol 136: 85-96. |
| [34] | Nunez L, Villalobos C, Boockfor FR, et al. (2000) The relationship between pulsatile secretion and calcium dynamics in single, living gonadotropin-releasing hormone neurons. Endocrinology 141: 2012-2017. |
| [35] | Sun W, Jarry H, Wuttke W, et al. (1997) Gonadotropin releasing hormone modulates gamma-aminobutyric acid-evoked intracellular calcium increase in immortalized hypothalamic gonadotropin releasing hormone neurons. Brain Res 747: 70-77. |
| [36] | Moore JP Jr, Shang E, Wray S (2002) In situ GABAergic modulation of synchronous gonadotropin releasing hormone-1 neuronal activity. J Neurosci 22: 8932-8941. |
| [37] | Terasawa E, Schanhofer WK, Keen KL, et al. (1999) Intracellular Ca2+ oscillations in luteinizing hormone-releasing hormone neurons derived from the embryonic olfactory placode of the rhesus monkey. J Neurosci 19: 5898-5909. |
| [38] | Richter TA, Keen KL, Terasawa E (2002) Synchronization of Ca2+ oscillations among primate LHRH neurons and nonneuronal cells in vitro. J Neurophysiol, 88: 1559-1567. |
| [39] | Silverman AJ, Witkin JW (1994) Biosynthesis of gonadotropin-releasing hormone during the rat estrous cycle: a cellular analysis. Neuroendocrinology 59: 545-551. |
| [40] | King JC, Tobet SA, Snavely FL, et al. (1982) LHRH immunopositive cells and their projections to the median eminence and organum vasculosum of the lamina terminalis. J Comp Neurol 209: 287-300. |
| [41] | Morelli A, Marini M, Mancina R, et al. (2008) Sex steroids and leptin regulate the "first Kiss" (KiSS 1/G-protein-coupled receptor 54 system) in human gonadotropin-releasing-hormone-secreting neuroblasts. J Sex Med 5: 1097-1113. |
| [42] | Morelli A, Fibbi B, Marini M, et al. (2009) Dihydrotestosterone and leptin regulate gonadotropin-releasing hormone (GnRH) expression and secretion in human GnRH-secreting neuroblasts. J Sex Med 6: 397-407. |
| [43] | Morelli A, Comeglio P, Sarchielli E, et al. (2013) Negative effects of high glucose exposure in human gonadotropin-releasing hormone neurons. Int J Endocrinol 2013: 684659. |
| [44] | Sarchielli E, Comeglio P, Squecco R, et al. (2016) Tumor Necrosis Factor alpha Impairs Kisspeptin Signaling in Human Gonadotropin-Releasing Hormone Primary Neurons. J Clin Endocrinol Metab: jc20162115. |
| [45] | Kimura F, Nishihara M, Hiruma H, et al. (1991) Naloxone increases the frequency of the electrical activity of luteinizing hormone-releasing hormone pulse generator in long-term ovariectomized rats. Neuroendocrinology 53: 97-102. |
| [46] | Nishihara M, Hiruma H, Kimura F (1991) Interactions between the noradrenergic and opioid peptidergic systems in controlling the electrical activity of luteinizing hormone-releasing hormone pulse generator in ovariectomized rats. Neuroendocrinology 54: 321-326. |
| [47] | Goubillon ML, Kaufman JM, Thalabard JC (1995) Hypothalamic multiunit activity and pulsatile luteinizing hormone release in the castrated male rat. Eur J Endocrinol 133: 585-590. |
| [48] | Mcgarvey C, Cates PA, Brooks A, et al. (2001) Phytoestrogens and gonadotropin-releasing hormone pulse generator activity and pituitary luteinizing hormone release in the rat. Endocrinology 142: 1202-1208. |
| [49] | Kinsey-Jones JS, Li XF, Luckman SM, et al. (2008) Effects of kisspeptin-10 on the electrophysiological manifestation of gonadotropin-releasing hormone pulse generator activity in the female rat. Endocrinology 149: 1004-1008. |
| [50] | Ohkura S, Tsukamura H, Maeda K (1991) Effects of various types of hypothalamic deafferentation on luteinizing hormone pulses in ovariectomized rats. J Neuroendocrinol 3: 503-508. |
| [51] | Jasoni CL, Todman MG, Strumia MM, et al. (2007) Cell type-specific expression of a genetically encoded calcium indicator reveals intrinsic calcium oscillations in adult gonadotropin-releasing hormone neurons. J Neurosci 27: 860-867. |
| [52] | Rodriguez EM, Blazquez JL, Guerra M (2010) The design of barriers in the hypothalamus allows the median eminence and the arcuate nucleus to enjoy private milieus: the former opens to the portal blood and the latter to the cerebrospinal fluid. Peptides 31: 757-776. |
| [53] | Terasawa E, Keen KL, Mogi K, et al. (1999) Pulsatile release of luteinizing hormone-releasing hormone (LHRH) in cultured LHRH neurons derived from the embryonic olfactory placode of the rhesus monkey. Endocrinology 140: 1432-1441. |
| [54] | Kuehl-Kovarik MC, Pouliot WA, Halterman GL, et al. (2002) Episodic bursting activity and response to excitatory amino acids in acutely dissociated gonadotropin-releasing hormone neurons genetically targeted with green fluorescent protein. J Neurosci 22: 2313-2322. |
| [55] | De Roux N, Genin E, Carel JC, et al. (2003) Hypogonadotropic hypogonadism due to loss of function of the KiSS1-derived peptide receptor GPR54. Proc Natl Acad Sci U S A 100: 10972-10976. |
| [56] | Seminara SB, Messager S, Chatzidaki EE, et al. (2003) The GPR54 gene as a regulator of puberty. N Engl J Med 349: 1614-1627. |
| [57] | Lee JH, Miele ME, Hicks DJ, et al. (1996) KiSS-1, a novel human malignant melanoma metastasis-suppressor gene. J Natl Cancer Inst 88: 1731-1737. |
| [58] | Kotani M, Detheux M, Vandenbogaerde A, et al. (2001) The metastasis suppressor gene KiSS-1 encodes kisspeptins, the natural ligands of the orphan G protein-coupled receptor GPR54. J Biol Chem 276: 34631-34636. |
| [59] | Muir AI, Chamberlain L, Elshourbagy NA, et al. (2001) AXOR12, a novel human G protein-coupled receptor, activated by the peptide KiSS-1. J Biol Chem 276: 28969-28975. |
| [60] | Ohtaki T, Shintani Y, Honda S, et al. (2001) Metastasis suppressor gene KiSS-1 encodes peptide ligand of a G-protein-coupled receptor. Nature 411: 613-617. |
| [61] | Kauffman AS (2010) Coming of age in the kisspeptin era: sex differences, development, and puberty. Mol Cell Endocrinol 324: 51-63. |
| [62] | Herbison AE (2008) Estrogen positive feedback to gonadotropin-releasing hormone (GnRH) neurons in the rodent: the case for the rostral periventricular area of the third ventricle (RP3V). Brain Res Rev 57: 277-287. |
| [63] | Smith JT, Li Q, Pereira A, et al. (2009) Kisspeptin neurons in the ovine arcuate nucleus and preoptic area are involved in the preovulatory luteinizing hormone surge. Endocrinology 150: 5530-5538. |
| [64] | Hrabovszky E, Ciofi P, Vida B, et al. (2010) The kisspeptin system of the human hypothalamus: sexual dimorphism and relationship with gonadotropin-releasing hormone and neurokinin B neurons. Eur J Neurosci 31: 1984-1998. |
| [65] | Lehman MN, Hileman SM, Goodman RL (2013) Neuroanatomy of the kisspeptin signaling system in mammals: comparative and developmental aspects. Adv Exp Med Biol 784: 27-62. |
| [66] | Goodman RL, Lehman MN, Smith JT, et al. (2007) Kisspeptin neurons in the arcuate nucleus of the ewe express both dynorphin A and neurokinin B. Endocrinology 148: 5752-5760. |
| [67] | Cheng G, Coolen LM, Padmanabhan V, et al. (2010) The kisspeptin/neurokinin B/dynorphin (KNDy) cell population of the arcuate nucleus: sex differences and effects of prenatal testosterone in sheep. Endocrinology 151: 301-311. |
| [68] | Li XF, Kinsey-Jones JS, Cheng Y, et al. (2009) Kisspeptin signalling in the hypothalamic arcuate nucleus regulates GnRH pulse generator frequency in the rat. PLoS One 4: e8334. |
| [69] | Irwig MS, Fraley GS, Smith JT, et al. (2004) Kisspeptin activation of gonadotropin releasing hormone neurons and regulation of KiSS-1 mRNA in the male rat. Neuroendocrinology 80: 264-272. |
| [70] | Navarro VM, Castellano JM, Fernandez-Fernandez R, et al. (2005) Effects of KiSS-1 peptide, the natural ligand of GPR54, on follicle-stimulating hormone secretion in the rat. Endocrinology 146: 1689-1697. |
| [71] | Navarro VM, Castellano JM, Fernandez-Fernandez R, et al. (2005) Characterization of the potent luteinizing hormone-releasing activity of KiSS-1 peptide, the natural ligand of GPR54. Endocrinology 146: 156-163. |
| [72] | Messager S, Chatzidaki EE, Ma D, et al. (2005) Kisspeptin directly stimulates gonadotropin-releasing hormone release via G protein-coupled receptor 54. Proc Natl Acad Sci U S A 102: 1761-1766. |
| [73] | Ohkura S, Takase K, Matsuyama S, et al. (2009) Gonadotrophin-releasing hormone pulse generator activity in the hypothalamus of the goat. J Neuroendocrinol 21: 813-821. |
| [74] | Plant TM (2006) The role of KiSS-1 in the regulation of puberty in higher primates. Eur J Endocrinol, 155 Suppl 1: S11-16. |
| [75] | Dhillo WS, Chaudhri OB, Thompson EL, et al. (2007) Kisspeptin-54 stimulates gonadotropin release most potently during the preovulatory phase of the menstrual cycle in women. J Clin Endocrinol Metab 92: 3958-3966. |
| [76] | Gottsch ML, Cunningham MJ, Smith JT, et al. (2004) A role for kisspeptins in the regulation of gonadotropin secretion in the mouse. Endocrinology 145: 4073-4077. |
| [77] | D'anglemont De Tassigny X, Fagg LA, Carlton MB, et al. (2008) Kisspeptin can stimulate gonadotropin-releasing hormone (GnRH) release by a direct action at GnRH nerve terminals. Endocrinology 149: 3926-3932. |
| [78] | Qiu J, Nestor CC, Zhang C, et al. (2016) High-frequency stimulation-induced peptide release synchronizes arcuate kisspeptin neurons and excites GnRH neurons. Elife 5. |
| [79] | Moenter SM (2010) Identified GnRH neuron electrophysiology: a decade of study. Brain Res 1364: 10-24. |
| [80] | Han SK, Gottsch ML, Lee KJ, et al. (2005) Activation of gonadotropin-releasing hormone neurons by kisspeptin as a neuroendocrine switch for the onset of puberty. J Neurosci 25: 11349-11356. |
| [81] | Pielecka-Fortuna J, Chu Z, Moenter SM (2008) Kisspeptin acts directly and indirectly to increase gonadotropin-releasing hormone neuron activity and its effects are modulated by estradiol. Endocrinology 149: 1979-1986. |
| [82] | Roseweir AK, Kauffman AS, Smith JT, et al. (2009) Discovery of potent kisspeptin antagonists delineate physiological mechanisms of gonadotropin regulation. J Neurosci 29: 3920-3929. |
| [83] | Choe HK, Kim HD, Park SH, et al. (2013) Synchronous activation of gonadotropin-releasing hormone gene transcription and secretion by pulsatile kisspeptin stimulation. Proc Natl Acad Sci U S A 110: 5677-5682. |
| [84] | Clarkson J, Herbison AE (2006) Postnatal development of kisspeptin neurons in mouse hypothalamus; sexual dimorphism and projections to gonadotropin-releasing hormone neurons. Endocrinology 147: 5817-5825. |
| [85] | Wintermantel TM, Campbell RE, Porteous R, et al. (2006) Definition of estrogen receptor pathway critical for estrogen positive feedback to gonadotropin-releasing hormone neurons and fertility. Neuron 52: 271-280. |
| [86] | Yeo SH, Herbison AE (2011) Projections of arcuate nucleus and rostral periventricular kisspeptin neurons in the adult female mouse brain. Endocrinology 152: 2387-2399. |
| [87] | Decourt C, Tillet Y, Caraty A, et al. (2008) Kisspeptin immunoreactive neurons in the equine hypothalamus Interactions with GnRH neuronal system. J Chem Neuroanat 36: 131-137. |
| [88] | Ramaswamy S, Guerriero KA, Gibbs RB, et al. (2008) Structural interactions between kisspeptin and GnRH neurons in the mediobasal hypothalamus of the male rhesus monkey (Macaca mulatta) as revealed by double immunofluorescence and confocal microscopy. Endocrinology 149: 4387-4395. |
| [89] | Roa J, Navarro VM, Tena-Sempere M (2011) Kisspeptins in reproductive biology: consensus knowledge and recent developments. Biol Reprod 85: 650-660. |
| [90] | Roa J, Castellano JM, Navarro VM, et al. (2009) Kisspeptins and the control of gonadotropin secretion in male and female rodents. Peptides 30: 57-66. |
| [91] | Goodman RL, Hileman SM, Nestor CC, et al. (2013) Kisspeptin, neurokinin B, and dynorphin act in the arcuate nucleus to control activity of the GnRH pulse generator in ewes. Endocrinology 154: 4259–4269. |
| [92] | Yip SH, Boehm U, Herbison AE, et al. (2015) Conditional Viral Tract Tracing Delineates the Projections of the Distinct Kisspeptin Neuron Populations to Gonadotropin-Releasing Hormone (GnRH) Neurons in the Mouse. Endocrinology 156: 2582-2594. |
| [93] | Smith JT, Dungan HM, Stoll EA, et al. (2005) Differential regulation of KiSS-1 mRNA expression by sex steroids in the brain of the male mouse. Endocrinology 146: 2976-2984. |
| [94] | Smith JT, Cunningham MJ, Rissman EF, et al. (2005) Regulation of Kiss1 gene expression in the brain of the female mouse. Endocrinology 146: 3686-3692. |
| [95] | Clarkson J, D'anglemont De Tassigny X, Moreno AS, et al. (2008) Kisspeptin-GPR54 signaling is essential for preovulatory gonadotropin-releasing hormone neuron activation and the luteinizing hormone surge. J Neurosci 28: 8691-8697. |
| [96] | Franceschini I, Lomet D, Cateau M, et al. (2006) Kisspeptin immunoreactive cells of the ovine preoptic area and arcuate nucleus co-express estrogen receptor alpha. Neurosci Lett 401: 225-230. |
| [97] | Smith JT (2013) Sex steroid regulation of kisspeptin circuits. Adv Exp Med Biol 784: 275-295. |
| [98] | Yamamura T, Wakabayashi Y, Sakamoto K, et al. (2014) The effects of chronic subcutaneous administration of an investigational kisspeptin analog, TAK-683, on gonadotropin-releasing hormone pulse generator activity in goats. Neuroendocrinology 100: 250-264. |
| [99] | Ramaswamy S, Seminara SB, Pohl CR, et al. (2007) Effect of continuous intravenous administration of human metastin 45-54 on the neuroendocrine activity of the hypothalamic-pituitary-testicular axis in the adult male rhesus monkey (Macaca mulatta). Endocrinology 148: 3364-3370. |
| [100] | Keen KL, Wegner FH, Bloom SR, et al. (2008) An increase in kisspeptin-54 release occurs with the pubertal increase in luteinizing hormone-releasing hormone-1 release in the stalk-median eminence of female rhesus monkeys in vivo. Endocrinology 149: 4151-4157. |
| [101] | Li Q, Millar RP, Clarke IJ, et al. (2015) Evidence that neurokinin B controls basal gonadotropin-releasing hormone secretion but is not critical for estrogen-positive feedback in sheep. Neuroendocrinology 101: 161-174. |
| [102] | Beale KE, Kinsey-Jones JS, Gardiner JV, et al. (2014) The physiological role of arcuate kisspeptin neurons in the control of reproductive function in female rats. Endocrinology 155: 1091-1098. |
| [103] | Hu MH, Li XF, Mccausland B, et al. (2015) Relative Importance of the Arcuate and Anteroventral Periventricular Kisspeptin Neurons in Control of Puberty and Reproductive Function in Female Rats. Endocrinology 156: 2619-2631. |
| [104] | Uenoyama Y, Nakamura S, Hayakawa Y, et al. (2015) Lack of pulse and surge modes and glutamatergic stimulation of luteinising hormone release in Kiss1 knockout rats. J Neuroendocrinol 27: 187-197. |
| [105] | Chan YM, Butler JP, Pinnell NE, et al. (2011) Kisspeptin resets the hypothalamic GnRH clock in men. J Clin Endocrinol Metab 96: E908-915. |
| [106] | George JT, Veldhuis JD, Roseweir AK, et al. (2011) Kisspeptin-10 is a potent stimulator of LH and increases pulse frequency in men. J Clin Endocrinol Metab 96: E1228-1236. |
| [107] | Chan YM, Butler JP, Sidhoum VF, et al. (2012) Kisspeptin administration to women: a window into endogenous kisspeptin secretion and GnRH responsiveness across the menstrual cycle. J Clin Endocrinol Metab 97: E1458-1467. |
| [108] | Hrabovszky E, Sipos MT, Molnar CS, et al. (2012) Low degree of overlap between kisspeptin, neurokinin B, and dynorphin immunoreactivities in the infundibular nucleus of young male human subjects challenges the KNDy neuron concept. Endocrinology 153: 4978-4989. |
| [109] | Young J, George JT, Tello JA, et al. (2013) Kisspeptin restores pulsatile LH secretion in patients with neurokinin B signaling deficiencies: physiological, pathophysiological and therapeutic implications. Neuroendocrinology 97: 193-202. |
| [110] | Fu LY, Van Den Pol AN (2010) Kisspeptin directly excites anorexigenic proopiomelanocortin neurons but inhibits orexigenic neuropeptide Y cells by an indirect synaptic mechanism. J Neurosci 30: 10205-10219. |
| [111] | Iijima N, Takumi K, Matsumoto K, et al. (2015) Visualization of Kisspeptin Binding to Rat Hypothalamic Neurons. Acta Histochem Cytochem 48: 179-184. |
| [112] | Guran T, Tolhurst G, Bereket A, et al. (2009) Hypogonadotropic hypogonadism due to a novel missense mutation in the first extracellular loop of the neurokinin B receptor. J Clin Endocrinol Metab 94: 3633-3639. |
| [113] | Topaloglu AK, Reimann F, Guclu M, et al. (2009) TAC3 and TACR3 mutations in familial hypogonadotropic hypogonadism reveal a key role for Neurokinin B in the central control of reproduction. Nat Genet 41: 354-358. |
| [114] | Burke MC, Letts PA, Krajewski SJ, et al. (2006) Coexpression of dynorphin and neurokinin B immunoreactivity in the rat hypothalamus: Morphologic evidence of interrelated function within the arcuate nucleus. J Comp Neurol 498: 712-726. |
| [115] | Amstalden M, Coolen LM, Hemmerle AM, et al. (2010) Neurokinin 3 receptor immunoreactivity in the septal region, preoptic area and hypothalamus of the female sheep: colocalisation in neurokinin B cells of the arcuate nucleus but not in gonadotrophin-releasing hormone neurones. J Neuroendocrinol 22: 1-12. |
| [116] | Navarro VM, Castellano JM, Mcconkey SM, et al. (2011) Interactions between kisspeptin and neurokinin B in the control of GnRH secretion in the female rat. Am J Physiol Endocrinol Metab 300: E202-210. |
| [117] | Grachev P, Porter KL, Coolen LM, et al. (2016) Surge-Like Luteinising Hormone Secretion Induced by Retrochiasmatic Area NK3R Activation is Mediated Primarily by Arcuate Kisspeptin Neurones in the Ewe. J Neuroendocrinol 28. |
| [118] | Sakamoto K, Murata K, Wakabayashi Y, et al. (2012) Central administration of neurokinin B activates kisspeptin/NKB neurons in the arcuate nucleus and stimulates luteinizing hormone secretion in ewes during the non-breeding season. J Reprod Dev 58: 700-706. |
| [119] | Ruka KA, Burger LL, Moenter SM (2016) Both Estrogen and Androgen Modify the Response to Activation of Neurokinin-3 and kappa-Opioid Receptors in Arcuate Kisspeptin Neurons From Male Mice. Endocrinology 157: 752-763. |
| [120] | Ruka KA, Burger LL, Moenter SM (2013) Regulation of arcuate neurons coexpressing kisspeptin, neurokinin B, and dynorphin by modulators of neurokinin 3 and kappa-opioid receptors in adult male mice. Endocrinology 154: 2761-2771. |
| [121] | Dellovade TL, Merchenthaler I (2004) Estrogen regulation of neurokinin B gene expression in the mouse arcuate nucleus is mediated by estrogen receptor alpha. Endocrinology 145: 736-742. |
| [122] | Navarro VM, Gottsch ML, Chavkin C, et al. (2009) Regulation of gonadotropin-releasing hormone secretion by kisspeptin/dynorphin/neurokinin B neurons in the arcuate nucleus of the mouse. J Neurosci 29: 11859-11866. |
| [123] | Grachev P, Millar RP, O’byrne KT (2014) The role of neurokinin B signalling in reproductive neuroendocrinology. Neuroendocrinology 99: 7-17. |
| [124] | Billings HJ, Connors JM, Altman SN, et al. (2010) Neurokinin B acts via the neurokinin-3 receptor in the retrochiasmatic area to stimulate luteinizing hormone secretion in sheep. Endocrinology 151: 3836-3846. |
| [125] | Wakabayashi Y, Nakada T, Murata K, et al. (2010) Neurokinin B and dynorphin A in kisspeptin neurons of the arcuate nucleus participate in generation of periodic oscillation of neural activity driving pulsatile gonadotropin-releasing hormone secretion in the goat. J Neurosci 30: 3124-3132. |
| [126] | Wakabayashi Y, Yamamura T, Sakamoto K, et al. (2013) Electrophysiological and morphological evidence for synchronized GnRH pulse generator activity among Kisspeptin/neurokinin B/dynorphin A (KNDy) neurons in goats. J Reprod Dev 59: 40-48. |
| [127] | Okamura H, Tsukamura H, Ohkura S, et al. (2013) Kisspeptin and GnRH pulse generation. Adv Exp Med Biol 784: 297-323. |
| [128] | Li SY, Li XF, Hu MH, et al. (2014) Neurokinin B receptor antagonism decreases luteinising hormone pulse frequency and amplitude and delays puberty onset in the female rat. J Neuroendocrinol 26: 521-527. |
| [129] | Foradori CD, Amstalden M, Goodman RL, et al. (2006) Colocalisation of dynorphin a and neurokinin B immunoreactivity in the arcuate nucleus and median eminence of the sheep. J Neuroendocrinol 18: 534-541. |
| [130] | Wuster M, Schulz R, Herz A (1980) Opiate activity and receptor selectivity of dynorphin1-13 and related peptides. Neurosci Lett 20: 79-83. |
| [131] | Gilbeau PM, Hosobuchi Y, Lee NM (1986) Dynorphin effects on plasma concentrations of anterior pituitary hormones in the nonhuman primate. J Pharmacol Exp Ther 238: 974-977. |
| [132] | Grachev P, Li XF, Kinsey-Jones JS, et al. (2012) Suppression of the GnRH pulse generator by neurokinin B involves a kappa-opioid receptor-dependent mechanism. Endocrinology 153: 4894-4904. |
| [133] | Goodman RL, Coolen LM, Anderson GM, et al. (2004) Evidence that dynorphin plays a major role in mediating progesterone negative feedback on gonadotropin-releasing hormone neurons in sheep. Endocrinology 145: 2959-2967. |
| [134] | Nakahara T, Uenoyama Y, Iwase A, et al. (2013) Chronic peripheral administration of kappa-opioid receptor antagonist advances puberty onset associated with acceleration of pulsatile luteinizing hormone secretion in female rats. J Reprod Dev 59: 479-484. |
| [135] | Goubillon ML, Forsdike RA, Robinson JE, et al. (2000) Identification of neurokinin B-expressing neurons as an highly estrogen-receptive, sexually dimorphic cell group in the ovine arcuate nucleus. Endocrinology 141: 4218-4225. |
| [136] | Mostari P, Ieda N, Deura C, et al. (2013) dynorphin-kappa opioid receptor signaling partly mediates estrogen negative feedback effect on LH pulses in female rats. J Reprod Dev 59: 266-272. |
| [137] | Foradori CD, Goodman RL, Adams VL, et al. (2005) Progesterone increases dynorphin a concentrations in cerebrospinal fluid and preprodynorphin messenger ribonucleic Acid levels in a subset of dynorphin neurons in the sheep. Endocrinology 146: 1835-1842. |
| [138] | Gottsch ML, Navarro VM, Zhao Z, et al. (2009) Regulation of Kiss1 and dynorphin gene expression in the murine brain by classical and nonclassical estrogen receptor pathways. J Neurosci 29: 9390-9395. |
| [139] | Ruiz-Pino F, Garcia-Galiano D, Manfredi-Lozano M, et al. (2015) Effects and interactions of tachykinins and dynorphin on FSH and LH secretion in developing and adult rats. Endocrinology 156: 576-588. |
| [140] | Weems PW, Witty CF, Amstalden M, et al. (2016) kappa-Opioid receptor is colocalized in GnRH and KNDy cells in the female ovine and rat brain. Endocrinology 157: 2367-2379. |
| [141] | Krajewski SJ, Anderson MJ, Iles-Shih L, et al. (2005) Morphologic evidence that neurokinin B modulates gonadotropin-releasing hormone secretion via neurokinin 3 receptors in the rat median eminence. J Comp Neurol 489: 372-386. |
| [142] | Krajewski SJ, Burke MC, Anderson MJ, et al. (2010) Forebrain projections of arcuate neurokinin B neurons demonstrated by anterograde tract-tracing and monosodium glutamate lesions in the rat. Neuroscience 166: 680-697. |
| [143] | Smith JT, Li Q, Yap KS, et al. (2011) Kisspeptin is essential for the full preovulatory LH surge and stimulates GnRH release from the isolated ovine median eminence. Endocrinology 152: 1001-1012. |
| [144] | Ramaswamy S, Seminara SB, Ali B, et al. (2010) Neurokinin B stimulates GnRH release in the male monkey (Macaca mulatta) and is colocalized with kisspeptin in the arcuate nucleus. Endocrinology 151: 4494-4503. |
| [145] | Merkley CM, Coolen LM, Goodman RL, et al. (2015) Evidence for changes in numbers of synaptic inputs onto KNDy and GnRH neurones during the preovulatory LH surge in the ewe. J Neuroendocrinol 27: 624-635. |
| [146] | Han SY, Mclennan T, Czieselsky K, et al. (2015) Selective optogenetic activation of arcuate kisspeptin neurons generates pulsatile luteinizing hormone secretion. Proc Natl Acad Sci U S A 112: 13109-13114. |
| [147] | Grachev P, Li XF, Lin YS, et al. (2012) GPR54-dependent stimulation of luteinizing hormone secretion by neurokinin B in prepubertal rats. PLoS One 7: e44344. |
| [148] | Ramaswamy S, Seminara SB, Plant TM (2011) Evidence from the agonadal juvenile male rhesus monkey (Macaca mulatta) for the view that the action of neurokinin B to trigger gonadotropin-releasing hormone release is upstream from the kisspeptin receptor. Neuroendocrinology 94: 237-245. |
| [149] | Garcia-Galiano D, Van Ingen Schenau D, Leon S, et al. (2012) Kisspeptin signaling is indispensable for neurokinin B, but not glutamate, stimulation of gonadotropin secretion in mice. Endocrinology 153: 316-328. |
| [150] | Narayanaswamy S, Prague JK, Jayasena CN, et al. (2016) Investigating the KNDy hypothesis in humans by co-administration of kisspeptin, neurokinin B and naltrexone in men. J Clin Endocrinol Metab: jc20161911. |
| [151] | Lehman MN, Coolen LM, Goodman RL (2010) Minireview: kisspeptin/neurokinin B/dynorphin (KNDy) cells of the arcuate nucleus: a central node in the control of gonadotropin-releasing hormone secretion. Endocrinology 151: 3479-3489. |
| [152] | Goodman RL, Parfitt DB, Evans NP, et al. (1995) Endogenous opioid peptides control the amplitude and shape of gonadotropin-releasing hormone pulses in the ewe. Endocrinology 136: 2412-2420. |
| [153] | Murakawa H, Iwata K, Takeshita T, et al. (2016) Immunoelectron microscopic observation of the subcellular localization of kisspeptin, neurokinin B and dynorphin A in KNDy neurons in the arcuate nucleus of the female rat. Neurosci Lett 612: 161-166. |
| [154] | Vaaga CE, Borisovska M, Westbrook GL (2014) Dual-transmitter neurons: functional implications of co-release and co-transmission. Curr Opin Neurobiol 29: 25-32. |
| [155] | Ferguson SS (2001) Evolving concepts in G protein-coupled receptor endocytosis: the role in receptor desensitization and signaling. Pharmacol Rev 53: 1-24. |
| [156] | Walther C, Ferguson SS (2013) Arrestins: role in the desensitization, sequestration, and vesicular trafficking of G protein-coupled receptors. Prog Mol Biol Transl Sci 118: 93-113. |
| [157] | Brown CH, Scott V, Ludwig M, et al. (2007) Somatodendritic dynorphin release: orchestrating activity patterns of vasopressin neurons. Biochem Soc Trans 35: 1236-1242. |
| [158] | Shuster SJ, Riedl M, Li X, et al. (2000) The kappa opioid receptor and dynorphin co-localize in vasopressin magnocellular neurosecretory neurons in guinea-pig hypothalamus. Neuroscience 96: 373-383. |
| [159] | Whitnall MH, Gainer H, Cox BM, et al. (1983) Dynorphin-A-(1-8) is contained within vasopressin neurosecretory vesicles in rat pituitary. Science 222: 1137-1139. |
| [160] | Shuster SJ, Riedl M, Li X, et al. (1999) Stimulus-dependent translocation of kappa opioid receptors to the plasma membrane. J Neurosci 19: 2658-2664. |
| [161] | Brown CH, Bourque CW (2006) Mechanisms of rhythmogenesis: insights from hypothalamic vasopressin neurons. Trends Neurosci 29: 108-115. |
| [162] | Brown CH, Bourque CW (2004) Autocrine feedback inhibition of plateau potentials terminates phasic bursts in magnocellular neurosecretory cells of the rat supraoptic nucleus. J Physiol 557: 949-960. |
| [163] | Weems PW, Coolen LM, Hileman SM, et al. (2016) Kappa opioid receptors are internalized in arcuate KNDy cells during GnRH pulse termination in the ewe. Program No. 60.04 / JJ16. 2016 Neuroscience Meeting Planner, 2016 San Diego, CA: . Society for Neuroscience. |
| [164] | Popa SM, Clifton DK, Steiner RA (2008) The role of kisspeptins and GPR54 in the neuroendocrine regulation of reproduction. Annu Rev Physiol 70: 213-238. |
| [165] | Mittelman-Smith MA, Krajewski-Hall SJ, Mcmullen NT, et al. (2016) Ablation of KNDy Neurons Results in Hypogonadotropic Hypogonadism and Amplifies the Steroid-Induced LH Surge in Female Rats. Endocrinology 157: 2015-2027. |
| [166] | Smith JT (2008) Kisspeptin signalling in the brain: steroid regulation in the rodent and ewe. Brain Res Rev 57: 288-298. |
| [167] | Mayer C, Acosta-Martinez M, Dubois SL, et al. (2010) Timing and completion of puberty in female mice depend on estrogen receptor alpha-signaling in kisspeptin neurons. Proc Natl Acad Sci U S A 107: 22693-22698. |
| [168] | De Croft S, Piet R, Mayer C, et al. (2012) Spontaneous kisspeptin neuron firing in the adult mouse reveals marked sex and brain region differences but no support for a direct role in negative feedback. Endocrinology 153: 5384-5393. |
| [169] | Dubois SL, Acosta-Martinez M, Dejoseph MR, et al. (2015) Positive, but not negative feedback actions of estradiol in adult female mice require estrogen receptor alpha in kisspeptin neurons. Endocrinology 156: 1111-1120. |
| [170] | Goodman RL, Inskeep EK (2015) Chapter 27 - Control of the Ovarian Cycle of the Sheep. In: Plant TM, Zeleznik AJ (eds.) Knobil and Neill's Physiology of Reproduction (Fourth Edition). San Diego: Academic Press. |
| [171] | Smith JT, Popa SM, Clifton DK, et al. (2006) Kiss1 neurons in the forebrain as central processors for generating the preovulatory luteinizing hormone surge. J Neurosci 26: 6687-6694. |
| [172] | Adachi S, Yamada S, Takatsu Y, et al. (2007) Involvement of anteroventral periventricular metastin/kisspeptin neurons in estrogen positive feedback action on luteinizing hormone release in female rats. J Reprod Dev 53: 367-378. |
| [173] | Merkley CM, Porter KL, Coolen LM, et al. (2012) KNDy (kisspeptin/neurokinin B/dynorphin) neurons are activated during both pulsatile and surge secretion of LH in the ewe. Endocrinology 153: 5406-5414. |
| [174] | Fergani C, Routly JE, Jones DN, et al. (2013) Kisspeptin, c-Fos and CRFR type 2 expression in the preoptic area and mediobasal hypothalamus during the follicular phase of intact ewes, and alteration after LPS. Physiol Behav 110-111: 158–168. |
| [175] | Hoffman GE, Le WW, Franceschini I, et al. (2011) Expression of fos and in vivo median eminence release of LHRH identifies an active role for preoptic area kisspeptin neurons in synchronized surges of LH and LHRH in the ewe. Endocrinology 152: 214-222. |
| [176] | Caraty A, Fabre-Nys C, Delaleu B, et al. (1998) Evidence that the mediobasal hypothalamus is the primary site of action of estradiol in inducing the preovulatory gonadotropin releasing hormone surge in the ewe. Endocrinology 139: 1752-1760. |
| [177] | Kinoshita M, Tsukamura H, Adachi S, et al. (2005) Involvement of central metastin in the regulation of preovulatory luteinizing hormone surge and estrous cyclicity in female rats. Endocrinology 146: 4431-4436. |
| [178] | Helena CV, Toporikova N, Kalil B, et al. (2015) KNDy Neurons Modulate the Magnitude of the Steroid-Induced Luteinizing Hormone Surges in Ovariectomized Rats. Endocrinology 156: 4200-4213. |
| [179] | Grachev P, Li XF, O'byrne K (2013) Stress regulation of kisspeptin in the modulation of reproductive function. Adv Exp Med Biol 784: 431-454. |
| [180] | Nestor CC, Kelly MJ, Ronnekleiv OK (2014) Cross-talk between reproduction and energy homeostasis: central impact of estrogens, leptin and kisspeptin signaling. Horm Mol Biol Clin Investig 17: 109-128. |
| [181] | Simonneaux V, Bur I, Ancel C, et al. (2012) A kiss for daily and seasonal reproduction. Prog Brain Res 199: 423-437. |
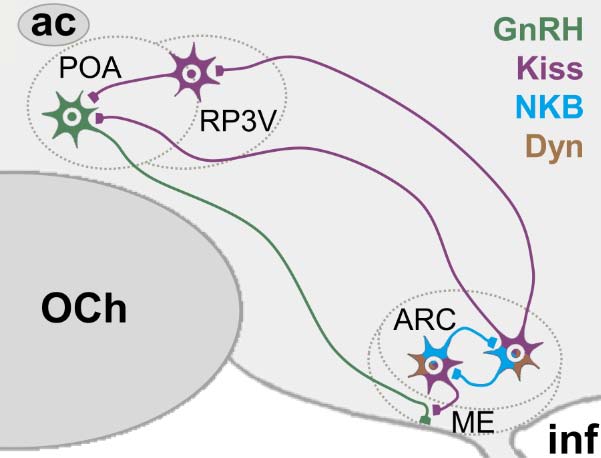
Figures(1)

Pasha Grachev, Robert L. Goodman. The GnRH Pulse Generator[J]. AIMS Medical Science, 2016, 3(4): 359-385. doi: 10.3934/medsci.2016.4.359







 DownLoad:
DownLoad: