Citation: Toshikazu Kuniya, Tarik Mohammed Touaoula. Global stability for a class of functional differential equations with distributed delay and non-monotone bistable nonlinearity[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 7332-7352. doi: 10.3934/mbe.2020375

| [1] | E. Braverman, D. Kinzebulatov, Nicholson's blowflies equation with distributed delay, Can. Appl. Math. Quart., 14 (2006), 107-128. |
| [2] |
C. Foley, M. C. Mackey, Dynamics hematological disease: a review, J. Math. Biol., 58 (2009), 285-322. doi: 10.1007/s00285-008-0165-3

|
| [3] | K. Gopalsamy, Stability and Oscillation in Delay Differential Equations of Population Dynamics, Springer, Dordrecht, 1992. |
| [4] |
W. S. C. Gurney, S. P. Blythe, R. M. Nisbet, Nicholson's blowflies revisited, Nature, 287 (1980), 17-21. doi: 10.1038/287017a0
|
| [5] |
C. Huang, Z. Yang, T. Yi, X. Zou, On the basins of attraction for a class of delay differential equations with non-monotone bistable nonlinearities, J. Diff. Equat., 256 (2014), 2101-2114. doi: 10.1016/j.jde.2013.12.015
|
| [6] | Y. Kuang, Delay Differential Equations: With Application in Population Dynamics, Academic Press, INC, 1993. |
| [7] |
M. C. Mackey, Unified hypothesis for the origin of aplastic anemia and periodic hematopoiesis, Blood, 51 (1978), 941-956. doi: 10.1182/blood.V51.5.941.941
|
| [8] |
E. Liz, A. Ruis-Herrera, Delayed population models with Allee effects and exploitation, Math. Biosci. Eng., 12 (2015), 83-97. doi: 10.3934/mbe.2015.12.83
|
| [9] |
E. Liz, M. Pinto, V. Tkachenko, S. Tromichuk, A global stability criterion for a family of delayed population models, Quart. Appl. Math., 63 (2005), 56-70. doi: 10.1090/S0033-569X-05-00951-3
|
| [10] |
E. Liz, G. Röst, On the global attractor of delay differential equations with unimodal feedback, Disc. Cont. Dyn. Syst., 24 (2009), 1215-1224. doi: 10.3934/dcds.2009.24.1215
|
| [11] |
E. Liz, V. Tkachenko, S. Trofimchuk, Mackey-Glass type delay differential equations near the boundary of absolute stability, J. Math. Anal. Appl., 275 (2002), 747-760. doi: 10.1016/S0022-247X(02)00416-X
|
| [12] |
E. Liz, V. Tkachenko, S. Tromichuk, A global stability criterion for scalar functional differential equations, SIAM J. Math. Anal., 35 (2003), 596-622. doi: 10.1137/S0036141001399222
|
| [13] |
G. Röst, J. Wu, Domain-decomposition method for the global dynamics of delay differential equations with unimodal feedback, Proc. R. Soc. A, 463 (2007), 2655-2669. doi: 10.1098/rspa.2007.1890
|
| [14] |
Y. Yuan, J. Belair, Stability and Hopf bifurcation analysis for functional differential equation with distributed delay, SIAM J. Appl. Dyn. Syst., 10 (2011), 551-581. doi: 10.1137/100794493
|
| [15] |
Y. Yuan, X. Q. Zhao, Global stability for non monotone delay equations (with application to a model of blood cell production), J. Diff. Equat., 252 (2012), 2189-2209. doi: 10.1016/j.jde.2011.08.026
|
| [16] | N. Bessonov, G. Bocharov, T. M. Touaoula, S. Trofimchuk, V. Volpert, Delay reaction-diffusion equation for infection dynamics, Disc. Cont. Dyn. Syst. Series B, 24 (2019), 2073-2091. |
| [17] |
K. Deng, Y. Wu, On the diffusive Nicholso's blowflies equation with distributed delay, Appl. Math. Lett., 50 (2015) 126-132. doi: 10.1016/j.aml.2015.06.013
|
| [18] |
W. Hu, Y. Zhou, Stability of a nonlocal delayed reaction-diffusion equation with a non-monotone bistable nonlinearity, Appl. Math. Lett., 79 (2018), 20-26. doi: 10.1016/j.aml.2017.11.018
|
| [19] | R. H. Martin, H. L. Smith, Abstract functional differential equations and reaction-diffusion systems, Trans. Amer. Math. Soc., 321 (1990), 1-44. |
| [20] | R. H. Martin, H. L. Smith, Reaction-diffusion systems with time delay: monotonicity, invariance, comparison and convergence, J. Reine Angew. Math., 413 (1991), 1-35. |
| [21] |
T. M. Touaoula, Global dynamics for a class of reaction-diffusion equations with distributed delay and Neumann condition, Commun. Pur. Appl. Anal., 19 (2020), 2473-2490. doi: 10.3934/cpaa.2020108
|
| [22] | T. M. Touaoula, M. N. Frioui, N. Bessonov, V. Volpert, Dynamics of solutions of a reactiondiffusion equation with delayed inhibition, Disc. Cont. Dyn. Syst. Series S, 13 (2020), 2425-2442. |
| [23] |
E. Braverman, S. Zhukovskiy, Absolute and delay-dependent stability of equations with a distributed delay, Disc. Cont. Dyn. Syst., 32 (2012), 2041-2061. doi: 10.3934/dcds.2012.32.2041
|
| [24] |
L. Berezansky, E. Braverman, L. Idels, Nicholson's blowflies differential equations revisited: main results and open problems, Appl. Math. Model., 34 (2010), 1405-1417. doi: 10.1016/j.apm.2009.08.027
|
| [25] |
L. Berezansky, E. Braverman, L. Idels, Mackey-Glass model of hematopoiesis with non-monotone feedback: stability, oscillation and control, Appl. Math. Compt., 219 (2013), 6268-6283. doi: 10.1016/j.amc.2012.12.043
|
| [26] |
H. A. El-Morshedy, Global attractivity in a population model with nonlinear death rate and distributed delays, J. Math. Anal. Appl., 410 (2014), 642-658. doi: 10.1016/j.jmaa.2013.08.060
|
| [27] | I. Gyori, S. Trofimchuk, Global attractivity in x'(t) = -δx(t) + pf (x(t - h)), Dynam. Syst. Appl., 8 (1999), 197-210. |
| [28] | H. L. Smith, H. R. Thieme, Dynamical Systems and Population Persistence, AMS, Providence, 2011. |
| [29] | H. L. Smith, Monotone Dynamical Systems: An Introduction to the Theory of Competitive and Cooperative Systems, AMS, Providence, 1995. |
| [30] | J. Hale, S. M. Verduyn Lunel, Introduction to Functional Differential Equations, Springer-Verlag, New York, 1993. |
| [31] |
T. M. Touaoula, Global stability for a class of functional differential equations (Application to Nicholson's blowflies and Mackey-Glass models), Disc. Cont. Dyn. Syst., 38 (2018), 4391-4419. doi: 10.3934/dcds.2018191
|
| [32] | H. R. Thieme, Mathematics in Population Biology, Princeton University Press, Princeton, 2003. |
| [33] |
T. Yi, Q. Li, Unstable sets, heteroclinic orbits and generic quasi-convergence for essentially strongly order-preserving semiflows, Proc. Edinb. Math. Soc., 52 (2009), 797-807. doi: 10.1017/S0013091506000320
|
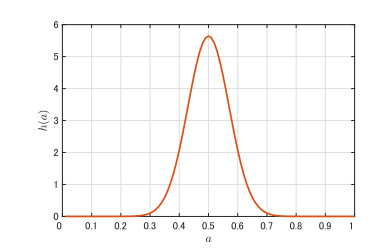
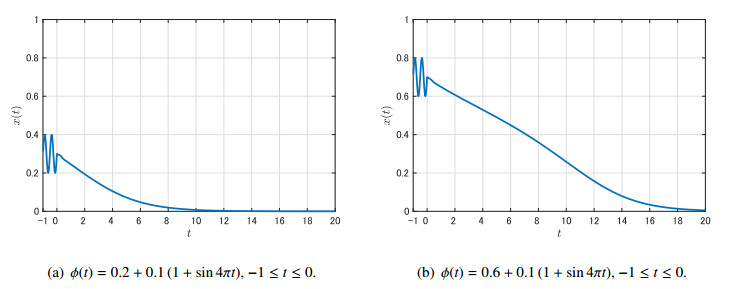
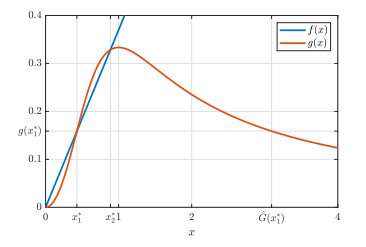
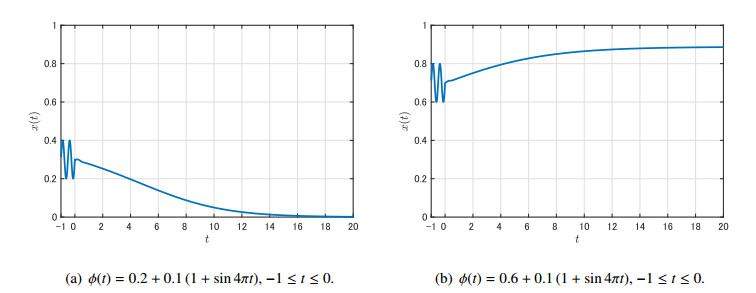
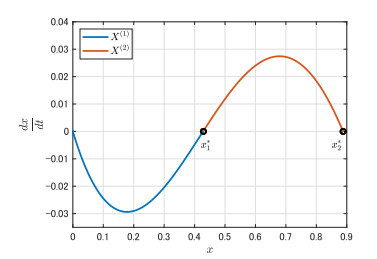
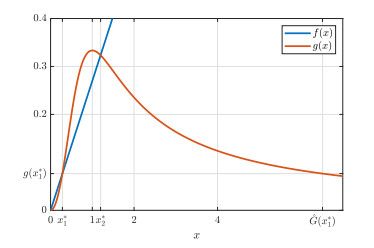
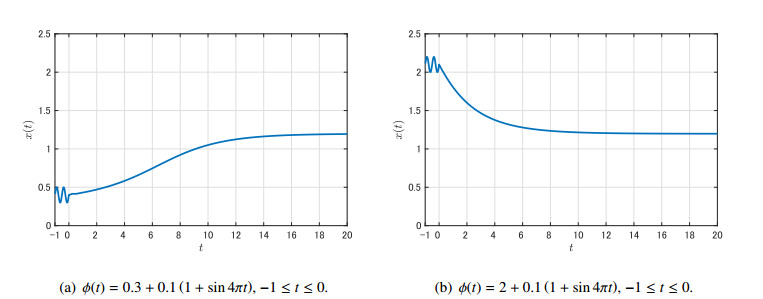
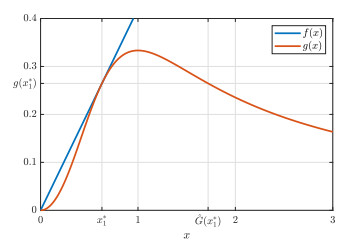
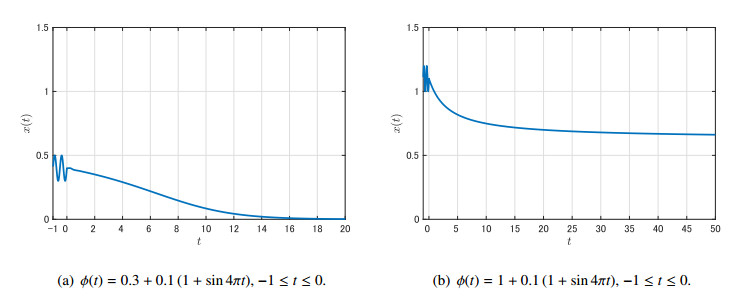
Figures(9)

Toshikazu Kuniya, Tarik Mohammed Touaoula. Global stability for a class of functional differential equations with distributed delay and non-monotone bistable nonlinearity[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 7332-7352. doi: 10.3934/mbe.2020375







 DownLoad:
DownLoad: