Citation: Meng Chen, Dacheng Jin, Bing Wang, Yunjiu Gou, Xinchun Dong. Identification of miRNAs as prognostic factors for esophageal squamous cell carcinoma[J]. Mathematical Biosciences and Engineering, 2020, 17(3): 2302-2309. doi: 10.3934/mbe.2020122

| [1] | W. Chen, R. Zheng, P. D. Baade, S. Zhang, H. Zeng, F. Bray, et al., Cancer statistics in China, 2015, Ca-Cancer J. Clin., 66 (2016), 115-132. |
| [2] | Z. Chen, J. Li, L. Tian, C. Zhou, Y. Gao, F. Zhou, et al., MiRNA expression profile reveals a prognostic signature for esophageal squamous cell carcinoma, Cancer Lett., 350 (2014), 34-42. |
| [3] | H. Z. Zhang, G. F. Jin, H. B. Shen, Epidemiologic differences in esophageal cancer between Asian and Western populations, Chin. J. Cancer, 2012, 31 (2012), 281. |
| [4] | Y. Guo, Z. Chen, L. Zhang, F. Zhou, S. Shi, X Feng, et al., Distinctive microRNA profiles relating to patient survival in esophageal squamous cell carcinoma, Cancer Res., 68 (2008), 26-33. |
| [5] | X. Wang, D. Kong, C. Wang, X. Ding, L. Zhang, M. Zhao, et al., Circulating microRNAs as novel potential diagnostic biomarkers for ovarian cancer: a systematic review and updated meta-analysis, J Ovarian Res., 12 (2019), 24. |
| [6] | C. W. Kan, V. M. Howell, M. A. Hahn, D. J. Marsh, Genomic alterations as mediators of miRNA dysregulation in ovarian cancer, Genes. Chromosomes Cancer, 54 (2015), 1-19. |
| [7] | X. H. Hu, J. Dai, H. L. Shang, Z. X. Zhao, Y. D. Hao, MiR-1285-3p is a potential prognostic marker in human osteosarcoma and functions as a tumor suppressor by targeting YAP1, Cancer Biomarkers, 25 (2019), 1-10. |
| [8] | J. von Frowein, S. M. Hauck, R. Kappler, P. Pagel, K. K. Fleischmann, T. Magg, et al., MiR‐492 regulates metastatic properties of hepatoblastoma via CD 44, Liver Int., 38 (2018), 1280-1291. |
| [9] | A. Fesler, H. Zhai, J. Ju, MiR-129 as a novel therapeutic target and biomarker in gastrointestinal cancer, OncoTargets Ther., 7 (2014), 1481. |
| [10] | A. Szpechcinski, M. Florczuk, K. Duk, A. Zdral, S. Rudzinski, M. Bryl, et al., The expression of circulating miR-504 in plasma is associated with EGFR mutation status in non-small-cell lung carcinoma patients, Cell. Mol. Life Sci., 2019, 1-16. |
| [11] | Y. Tan, J. J. Lin, X. Yang, D. M. Gou, L. Fu, F. R Li, et al., A panel of three plasma microRNAs for colorectal cancer diagnosis, Cancer Epidemiol., 60 (2019), 67-76. |
| [12] | L. L. Wong, R. Zou, L. Zhou, J. Y. Lim, D. C. Phua, C. Liu, et al., Combining Circulating MicroRNA and NT-proBNP to Detect and Categorize Heart Failure Subtypes, J. Am. Coll. Cardiol., 73 (2019), 1300-1313. |
| [13] | N. S. Altman, An introduction to kernel and nearest-neighbor nonparametric regression, Am. Stat., 46 (1992), 175-185. |
| [14] | T. H. Zhao, M. Jiang, T. Huang, B. Q. Li, N. Zhang, H. P. Li, et al, A novel method of predicting protein disordered regions based on sequence features, Biomed. Res. Int., 2013 (2013). |
| [15] | A. K. Rustgi, H. B. El-Serag, Esophageal carcinoma, New Engl. J Med., 371 (2014), 2499-2509. |
| [16] | Y. Song, L. Li, Y. Ou, Z. Gao, E. Li, X. Li, et al., Identification of genomic alterations in oesophageal squamous cell cancer, Nature, 509 (2014), 91. |
| [17] | K. J. Kaneko, M. L. DePamphilis, Regulation of gene expression at the beginning of mammalian development and the TEAD family of transcription factors, Dev. Genet., 22 (1998), 43-55. |
| [18] | Z. Shi, F. He, M. Chen, L. Hua, W. Wang, S. Jiao, et al., DNA-binding mechanism of the Hippo pathway transcription factor TEAD4, Oncogene, 36 (2017), 4362. |
| [19] | N. Nishioka, S. Yamamoto, H. Kiyonari, H. Sato, A. Sawada, M. Ota, et al., Tead4 is required for specification of trophectoderm in pre-implantation mouse embryos, Mech. Develop., 125 (2008), 270-283. |
| [20] | N. Nishioka, K. I. Inoue, K. Adachi, H. Kiyonari, M. Ota, A. Ralston, et al., The Hippo signaling pathway components Lats and Yap pattern Tead4 activity to distinguish mouse trophectoderm from inner cell mass, Dev. Cell, 16 (2009), 398-410. |
| [21] |
S. Takeuchi, A. Kasamatsu, M. Yamatoji, D. Nakashima, Y. Endo-Sakamoto, N. Koide, et al., TEAD4-YAP interaction regulates tumoral growth by controlling cell-cycle arrest at the G1 phase, Biochem. Biophys. Res. Commun., 486 (2017), 385-390. doi: 10.1016/j.bbrc.2017.03.050

|
| [22] | Y. Liu, G. Wang, Y. Yang, Z. Mei, Z. Liang, A. Cui, et al., Increased TEAD4 expression and nuclear localization in colorectal cancer promote epithelial-mesenchymal transition and metastasis in a YAP-independent manner, Oncogene, 35 (2016), 2789. |
| [23] | Q. Zhang, H. Fan, Q. Zou, H. Liu, B. Wan, S. Zhu, et al., TEAD 4 exerts pro‐metastatic effects and is negatively regulated by miR6839‐3p in lung adenocarcinoma progression, J Cell. Mol. Med., 22 (2018), 3560-3571. |
| [24] | S. M. Wang, W. X. Zeng, W. S. Wu, L. L. Sun, D. Yan,. Association between a microRNA binding site polymorphism in SLCO1A2 and the risk of delayed methotrexate elimination in Chinese children with acute lymphoblastic leukemia, Leukemia Res., 65 (2018), 61-66. |
| [25] | R. Fang, Y. Zhu, L. Hu, J. Ai, H. Zou, D. Ju, et al., Plasma MicroRNA Pair Panels as Novel Biomarkers for Detection of Early Stage Breast Cancer, Front. Physiol., 9, (2018),1879. |
| [26] | M. A.Chaudhry, Radiation‐induced microRNA: Discovery, functional analysis, and cancer radiotherapy, J Cell. Biochem., 115 (2014), 436-449. |
| [27] | E. van Schooneveld, H. Wildiers, I. Vergote, P. B. Vermeulen, L. Y. Dirix, et al., Dysregulation of microRNAs in breast cancer and their potential role as prognostic and predictive biomarkers in patient management, Breast Cancer Res., 17 (2015), 21. |
| [28] | J. Li, X. Kong, J. Zhang, Q. Luo, X. Li, L. Fang, MiRNA-26b inhibits proliferation by targeting PTGS2 in breast cancer, Cancer Cell Int., 13 (2013), 7. |
| [29] | C. M. Wilke, J. Hess, S. V. Klymenko, V. V. Chumak, L. M. Zakhartseva, E. V. Bakhanova, et al., Expression of mi RNA‐26b‐5p and its target TRPS 1 is associated with radiation exposure in post‐C hernobyl breast cancer, Int. J Cancer, 142 (2018), 573-583. |
| [30] | T. Liu, X. You, J. Sui, B. Shen, Y. Zhang, X. M. Zhang, et al., Prognostic value of a two‐microRNA signature for papillary thyroid cancer and a bioinformatic analysis of their possible functions, J Cell. Biochem., 120 (2019), 7185-7198. |
| [31] | M. L. Slattery, J. S. Herrick, L. E. Mullany, N. Valeri, J. Stevens, B. J. Caan, et al., An evaluation and replication of mi RNA s with disease stage and colorectal cancer‐specific mortality, Int. J Cancer, 137 (2015), 428-438. |
| [32] | L. Falzone, L. Scola, A. Zanghì, A. Biondi, A. D. Cataldo, M. Libra, et al., Integrated analysis of colorectal cancer microRNA datasets: Identification of microRNAs associated with tumor development, Aging, 10 (2018), 1000. |
| [33] | L. Falzone, L. Scola, A. Zanghì, A. Biondi, A. Di Cataldo, M. Libra, et al., MicroRNA-125b-5p regulates IL-1β induced inflammatory genes via targeting TRAF6-mediated MAPKs and NF-κB signaling in human osteoarthritic chondrocytes, Sci. Rep., 9 (2019), 6882. |
| [34] | A. Zeybek, N. Öz, S. Kalemci, T. Edgünlü, M. T. Kızıltuğ, K. Tosun, et al., Diagnostic Value of MiR-125b as a Potential Biomarker for Stage I Lung Adenocarcinoma Lung Adenocarcinoma, Curr. Mol. Med., 19 (2019), 216-227. |
| [35] | C. Martínez, B. K. Rodiño-Janeiro, B. Lobo, M. L. Stanifer, B. Klaus, M. Granzow, et al., MiR-16 and miR-125b are involved in barrier function dysregulation through the modulation of claudin-2 and cingulin expression in the jejunum in IBS with diarrhea, Gut, 66 (2017), 1537-1538. |
| [36] | X. Yang, C. Zhang, H. Tie, J. Luo, Y. Wang, Q. Wu, MiR-760 exerts an antioncogenic effect in esophageal squamous cell carcinoma by negatively driving fat metabolism via targeting c-Myc, J Cell. Biochem., 2019. |
| [37] | J. Tang, Z. Li, Q. Zhu, W. Wen, J. Wang, J. Xu, et al., MiR‐204‐5p regulates cell proliferation, invasion, and apoptosis by targeting IL‐11 in esophageal squamous cell carcinoma, J Cell. Physiol., 2019. |
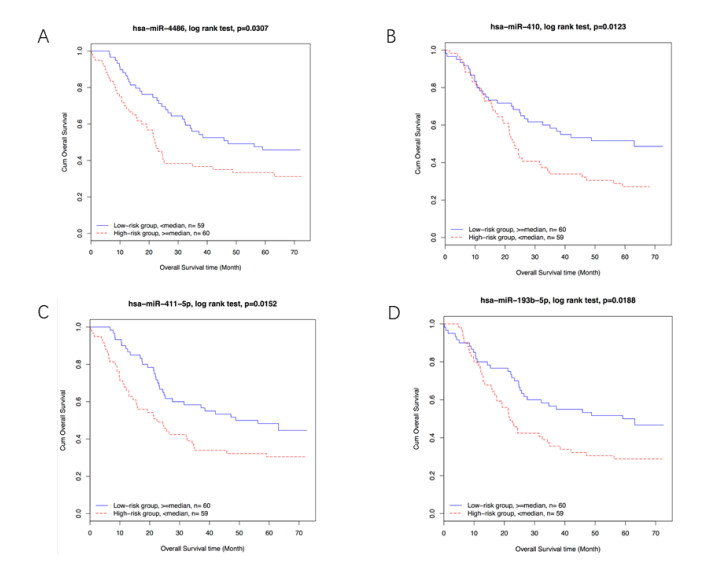
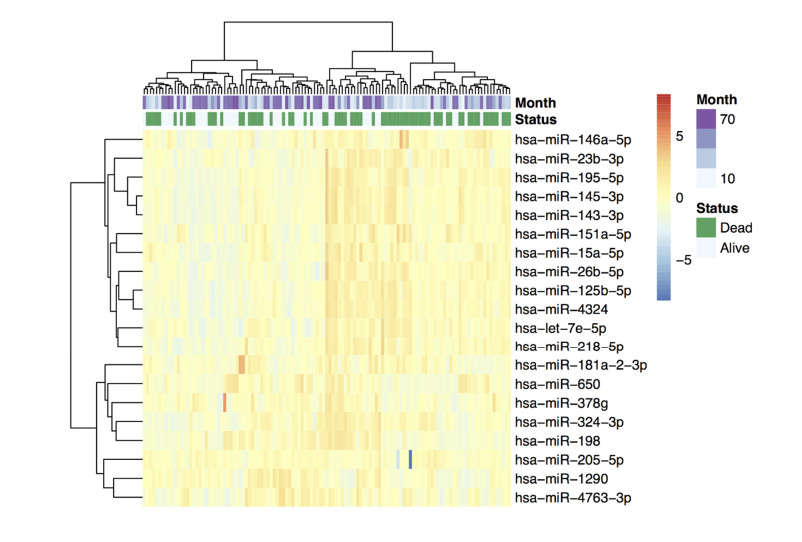
Figures(2)

Meng Chen, Dacheng Jin, Bing Wang, Yunjiu Gou, Xinchun Dong. Identification of miRNAs as prognostic factors for esophageal squamous cell carcinoma[J]. Mathematical Biosciences and Engineering, 2020, 17(3): 2302-2309. doi: 10.3934/mbe.2020122







 DownLoad:
DownLoad: