Citation: Xuejuan Lu, Shaokai Wang, Shengqiang Liu, Jia Li. An SEI infection model incorporating media impact[J]. Mathematical Biosciences and Engineering, 2017, 14(5&6): 1317-1335. doi: 10.3934/mbe.2017068

| [1] | [ M. Becker,J. Joseph, AIDS and behavioral change to reduce risk: A review, Amer. J. Public Health, 78 (1988): 394-410. |
| [2] | [ R. J. Blendon,J. M. Benson,C. M. DesRoches,E. Raleigh,K. Taylor-Clark, The public's response to severe acute respiratory syndormw in Toronto and the United States, Clinic Infectious Diseases, 38 (2004): 925-931. |
| [3] | [ S. M. Blower,A. R. McLean, Prophylactic vaccines, risk behavior change, and the probability of eradicating HIV in San Francisco, Science, 265 (1994): 1451-1454. |
| [4] | [ F. Brauer and C. Castillo-Chavez, Mathematical Models in Population Biology and Epidemics, Springer-Verlag, New York, 2001. |
| [5] | [ S. Busenberg and K. Cooke, Vertically Transmitted Diseases, Springer-Verlag, New York, 1993. |
| [6] | [ L. Cai,X. Li, Analysis of a SEIV epidemic model with a nonlinear incidence rate, Applied Mathematical Modelling, 33 (2009): 2919-2926. |
| [7] | [ V. Capasso,G. Serio, A generalisation of the Kermack-McKendrick deterministic epidemic model, Math. Biosci., 42 (1978): 43-61. |
| [8] | [ R. Chunara,J. R. Andrews,J. S. Brownstein, Social and news media enable estimation of epidemiological patterns early in the 2010 Haitian cholera outbreak, Am. J. Trop. Med. Hyg., 86 (2012): 39-45. |
| [9] | [ S. Collinson and J. M. Heffernan, Modelling the effects of media during an influenza epidemic, BMC Public Health, 14 (2014), 376. |
| [10] | [ J. Cui,Y. Sun,H. Zhu, The impact of media on the control of infectious diseases, Journal of Dynamics and Differential Equations, 20 (2008): 31-53. |
| [11] | [ J. Cui,X. Tao,H. Zhu, An SIS infection model incorporating media coverage, Rocky Mountain Journal of Mathematics, 38 (2008): 1323-1334. |
| [12] | [ O. Diekmann and J. Heesterbeek, Mathematical Epidemiology of Infectious Diseases: Model Building, Analysis and Interpretation, Wiley, New York, 2000. |
| [13] | [ K. A. Fanti,E. Vanman,C. C. Henrich,M. N. Avraamide, Desensitization to media violence over a short period of time, Aggressive Behavior, 35 (2009): 179-187. |
| [14] | [ K. Frost,E. Frank,E. Maibach, Relative risk in the news media: A quantification of misrepresentation, American Journal of Public Health, 87 (1997): 842-845. |
| [15] | [ H. Hethcote, The mathematics of infectious diseases, SIAM Revi., 42 (2000): 599-653. |
| [16] | [ X. Lai,S. Liu,R. Lin, Rich dynamical behaviors for predator-prey model with weak Allee effect, Applicable Analysis, 89 (2010): 1271-1292. |
| [17] | [ J. LaSalle, The Stability of Dynamical Systems, Reg. Conf. Ser. Appl. Math., SIAM, Philadelphia, 1976. |
| [18] | [ J. LaSalle and S. Lefschetz, Stability by Lyapunov's Direct Method, Academic Press, New York, 1961. |
| [19] | [ G. M. Leung, T. H. Lam, L. M. Ho, S. Y. Ho, B. H. Chang, I. O. Wong and A. J. Hedley, The impac of community sphychological response on outbreak control for severe acute respiratory syndrome in Hong Kong, J. Epid. Comm. Health 2003, p995. |
| [20] | [ S. Levin, T. Hallam and L. Gross, Applied Mathematical Ecology, Lecture Notes in Biomathematics, vol. 18, Springer, Berlin, Germany, 1989. |
| [21] | [ B. Li, S. Liu, J. Cui and J. Li, A simple predator-prey population model with rich dynamics, App. Sci., 6 (2016), 151. |
| [22] | [ R. Lin,S. Liu,X. Lai, Bifurcations of a Predator-prey System with Weak Allee effects, J. Korean Math. Soc., 50 (2013): 695-713. |
| [23] | [ R. Liu,J. Wu,H. Zhu, Media/psychological impact on multiple outbreaks of emerging infectious diseases, Computational and Mathematical Methods in Medicine, 8 (2007): 153-164. |
| [24] | [ W. Liu,H. Hethcote,S. Levin, Dynamical behaviour of epidemiological models with nonlinear incidence rates, J. Math. Biol., 25 (1987): 359-380. |
| [25] | [ W. Liu,S. Levin,Y. Iwasa, Influence of nonlinear incidence rates upon the behaviour of SIRS epidemiological models, J. Math. Biol., 23 (1986): 187-204. |
| [26] | [ Y. Liu,J. Cui, The impact of media coverage on the dynamics of infectious disease, International Journal of Biomathematics, 1 (2008): 65-74. |
| [27] | [ A. K. Misra,A. Sharma,J. Li, A mathematical model for control of vector borne diseases through media campaigns, Discret. Contin. Dyn. S., 18 (2013): 1909-1927. |
| [28] | [ A. K. Misra,A. Sharma,J. B. Shukla, Modeling and analysis of effects of awareness programs by media on the spread of infectious diseases, Math. Comput. Model, 53 (2011): 1221-1228. |
| [29] | [ A. K. Misra,A. Sharma,V. Singh, Effect of awareness programs in controlling the prevalence of an epidemic with time delay, J. Biol. Sys., 19 (2011): 389-402. |
| [30] | [ A. Mummert and H. Weiss, Get the news out loudly and quickly: The Influence of the media on limiting emerging infectious disease outbreaks, PloS One, 8 (2013), e71692. |
| [31] | [ J. Murray, Mathematical Biology, Springer-Verlag, Berlin, 1989. |
| [32] | [ C. Sun,W. Yang,J. Arino,K. Khan, Effect of media-induced social distancing on disease transmission in a two patch setting, Math. Biosci., 230 (2011): 87-95. |
| [33] | [ J. Tchuenche, N. Dube, C. Bhunu, R. Smith and C. Bauch, The impact of media coverage on the transmission dynamics of human inuenza, BMC Public Health, 11 (2011), S5. |
| [34] | [ P. van den Driessche,J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of diseases transmission, Math. Biosci., 180 (2002): 29-48. |
| [35] | [ A. A. F. Wahlberg,L. Sjoberg, Risk perception and the media, Journal of Risk Research, 3 (2001): 31-50. |
| [36] | [ Y. Xiao, S. Tang and J. Wu, Media impact switching surface during an infectious disease outbreak, Scientific Reports, 5 (2015), 7838. |
| [37] | [ Y. Xiao,T. Zhao,S. Tang, Dynamics of an infectious diseases with media/psychology induced non-smooth incidence, Math. Biosci. and Eng., 10 (2013): 445-461. |
| [38] | [ M. E. Young, G. R. Norman and K. R. Humphreys, Medicine in the popular press: The Influence of the media on perceptions of disease, PloS One, (3) (2008), e3552. |
| [39] | [ X. Zhao, Uniform persistence and periodic coexistence states in infinite-dimensional periodic semiflow with applications, Can. Appl. Math. Q., 3 (1995): 473-495. |

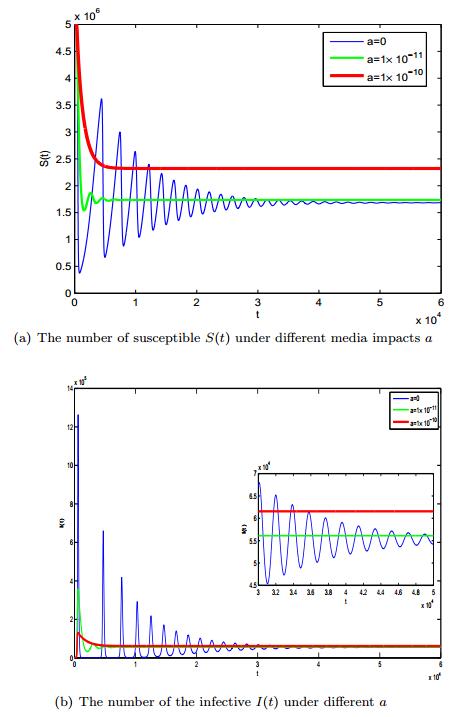
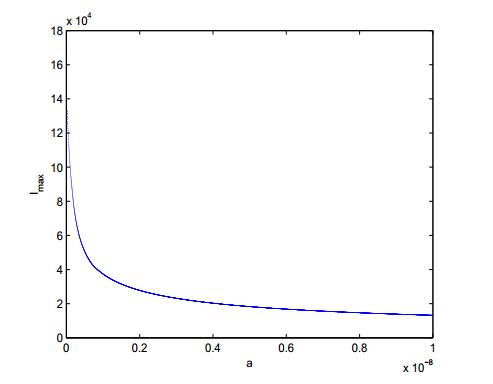
Figures(3) / Tables(1)

Xuejuan Lu, Shaokai Wang, Shengqiang Liu, Jia Li. An SEI infection model incorporating media impact[J]. Mathematical Biosciences and Engineering, 2017, 14(5&6): 1317-1335. doi: 10.3934/mbe.2017068







 DownLoad:
DownLoad: