Citation: Kyohei Fujita, Zhonggang Feng, Daisuke Sato, Tadashi Kosawada, Takao Nakamura, Yasuyuki Shiraishi, Mitsuo Umezu. Modulation of the mechanical properties of ventricular extracellular matrix hydrogels with a carbodiimide crosslinker and investigation of their cellular compatibility[J]. AIMS Materials Science, 2018, 5(1): 54-74. doi: 10.3934/matersci.2018.1.54

| [1] | Vunjak-Novakovic G, Tandon N, Godier A, et al. (2009) Challenges in cardiac tissue engineering. Tissue Eng Part B 16: 169–187. |
| [2] |
Leone M, Magadum A, Engel FB (2015) Cardiomyocyte proliferation in cardiac development and regeneration: a guide to methodologies and interpretations. Am J Physiol-Heart C 309: H1237–H1250. doi: 10.1152/ajpheart.00559.2015

|
| [3] |
Mummery CL, Zhang J, Ng ES, et al. (2012) Differentiation of human embryonic stem cells and induced pluripotent stem cells to cardiomyocytes: a method overview. Circ Res 111: 344–358. doi: 10.1161/CIRCRESAHA.110.227512
|
| [4] |
Schwach V, Passier R (2016) Generation and purification of human stem cell-derived cardiomyocytes. Differentiation 91: 126–138. doi: 10.1016/j.diff.2016.01.001
|
| [5] |
Kempf H, Andree B, Zweigerdt R (2016) Large-scale production of human pluripotent stem cell derived cardiomyocytes. Adv Drug Deliver Rev 96: 18–30. doi: 10.1016/j.addr.2015.11.016
|
| [6] |
Duan Y, Liu Z, O'Neill J, et al. (2011) Hybrid gel composed of native heart matrix and collagen induces cardiac differentiation of human embryonic stem cells without supplemental growth factors. J Cardiovasc Transl 4: 605–615. doi: 10.1007/s12265-011-9304-0
|
| [7] |
Higuchi S, Lin Q, Wang J, et al. (2013) Heart extracellular matrix supports cardiomyocyte differentiation of mouse embryonic stem cells. J Biosci Bioeng 115: 320–325. doi: 10.1016/j.jbiosc.2012.10.004
|
| [8] |
Engler AJ, Sen S, Sweeney HL, et al. (2006) Matrix elasticity directs stem cell lineage specification. Cell 126: 677–689. doi: 10.1016/j.cell.2006.06.044
|
| [9] |
Chaudhuri O, Gu L, Darnell M, et al. (2015) Substrate stress relaxation regulates cell spreading. Nat Commun 6: 6364. doi: 10.1038/ncomms7364
|
| [10] |
Brandl F, Sommer F, Goepferich A (2007) Rational design of hydrogels for tissue engineering: impact of physical factors on cell behavior. Biomaterials 28: 134–146. doi: 10.1016/j.biomaterials.2006.09.017
|
| [11] |
Feng Z, Wagatsuma Y, Kikuchi M, et al. (2014) The mechanisms of fibroblast-mediated compaction of collagen gels and the mechanical niche around individual fibroblasts. Biomaterials 35: 8078–8091. doi: 10.1016/j.biomaterials.2014.05.072
|
| [12] |
Feng Z, Ishiguro Y, Fujita K, et al. (2015) A fibril-based structural constitutive theory reveals the dominant role of network characteristics on the mechanical behavior of fibroblast-compacted collagen gels. Biomaterials 67: 365–381. doi: 10.1016/j.biomaterials.2015.07.038
|
| [13] |
Johnson TD, Lin SY, Christman KL (2011) Tailoring material properties of a nanofibrous extracellular matrix derived hydrogel. Nanotechnology 22: 494015. doi: 10.1088/0957-4484/22/49/494015
|
| [14] |
Freytes DO, Martin J, Velankar SS, et al. (2008) Preparation and rheological characterization of a gel form of the porcine urinary bladder matrix. Biomaterials 29: 1630–1637. doi: 10.1016/j.biomaterials.2007.12.014
|
| [15] |
Williams C, Budina E, Stoppel WL, et al. (2015) Cardiac Extracellular Matrix-Fibrin Hybrid Scaffolds with Tunable Properties for Cardiovascular Tissue Engineering. Acta Biomater 14: 84–95. doi: 10.1016/j.actbio.2014.11.035
|
| [16] |
Grover GN, Rao N, Christman KL (2014) Myocardial Matrix-Polyethylene Glycol Hybrid Hydrogels for Tissue Engineering. Nanotechnology 25: 014011. doi: 10.1088/0957-4484/25/1/014011
|
| [17] |
Singelyn JM, Christman KL (2011) Modulation of material properties of a decellularized myocardial matrix scaffold. Macromol Biosci 11: 731–738. doi: 10.1002/mabi.201000423
|
| [18] | Everaerts F, Torrianni M, Hendriks M, et al. (2008) Biomechanical properties of carbodiimide crosslinked collagen: Influence of the formation of ester crosslinks. J Biomed Mater Res A 85: 547–555. |
| [19] | Fujita K, Tuchida Y, Seki H, et al. (2015) Characterizing and modulating the mechanical properties of hydrogels from ventricular extracellular matrix. 2015 10th Asian Control Conference (ASCC) 1–5. |
| [20] | Kikuchi M, Feng Z, Kosawada T, et al. (2016) Stress relaxation and stress-strain characteristics of porcine amniotic membrane. Bio-med Mater Eng 27: 603–611. |
| [21] |
Johnson TD, DeQuach JA, Gaetani R, et al. (2014) Human versus porcine tissue sourcing for an injectable myocardial matrix hydrogel. Biomater Sci 2: 735–744. doi: 10.1039/C3BM60283D
|
| [22] |
Ishihama Y, Oda Y, Tabata T, et al. (2005) Exponentially Modified Protein Abundance Index (emPAI) for Estimation of Absolute Protein Amount in Proteomics by the Number of Sequenced Peptides per Protein. Mol Cell Proteomics 4: 1265–1272. doi: 10.1074/mcp.M500061-MCP200
|
| [23] | Fung YC (1993) Biomechanics: Mechanical properties of living tissues, 2 Eds, Springer. |
| [24] |
Fancey KS (2005) A mechanical model for creep, recovery and stress relaxation in polymeric materials. J Mater Sci 40: 4827–4831. doi: 10.1007/s10853-005-2020-x
|
| [25] |
Feng Z, Matsumoto T, Nakamura T, et al. (2003) Measurements of the mechanical properties of contracted collagen gels populated with rat fibroblasts or cardiomyocytes. J Artif Organs 6: 192–196. doi: 10.1007/s10047-003-0230-z
|
| [26] |
Ott HC, Matthiesen TS, Goh SK, et al. (2008) Perfusion-decellularized matrix: using nature's platform to engineer a bioartificial heart. Nat Med 14: 213–221. doi: 10.1038/nm1684
|
| [27] |
Eiselt P, Lee KY, Mooney DJ (1999) Rigidity of Two-Component Hydrogels Prepared from Alginate and Poly(ethylene glycol)-Diamines. Macromolecules 32: 5561–5566. doi: 10.1021/ma990514m
|
| [28] |
Lee KY, Rowley JA, Eiselt P, et al. (2000) Controlling Mechanical and Swelling Properties of Alginate Hydrogels Independently by Cross-Linker Type and Cross-Linking Density. Macromolecules 33: 4291–4294. doi: 10.1021/ma9921347
|
| [29] |
Wang Y, Wang G, Luo X, et al. (2012) Substrate stiffness regulated the proliferation, migration, and differentiation of epidermal cells. Burns 38: 414–420. doi: 10.1016/j.burns.2011.09.002
|
| [30] |
Ungerleider JL, Johnson TD, Rao N, et al. (2015) Fabrication and Characterization of Injectable Hydrogels Derived from Decellularized Skeletal and Cardiac Muscle. Methods 84: 53–59. doi: 10.1016/j.ymeth.2015.03.024
|
| [31] |
Johnson TD, Dequach JA, Gaetani R, et al. (2014) Human versus porcine tissue sourcing for an injectable myocardial matrix hydrogel. Biomater Sci 2: 735–744. doi: 10.1039/C3BM60283D
|
| [32] |
Seif-Naraghi SB, Horn D, Schup-Magoffin PA, et al. (2011) Patient-to-patient variability in autologous pericardial matrix scaffolds for cardiac repair. J Cardiovasc Transl 4: 545–556. doi: 10.1007/s12265-011-9293-z
|
| [33] |
Seif-Naraghi SB, Horn D, Schup-Magoffin PJ, et al. (2012) Injectable extracellular matrix derived hydrogel provides a platform for enhanced retention and delivery of a heparin-binding growth factor. Acta Biomater 8: 3695–3703. doi: 10.1016/j.actbio.2012.06.030
|
| [34] |
Huang-Lee LL, Cheung DT, Nimni ME (1990) Biochemical changes and cytotoxicity associated with the degradation of polymeric glutaraldehyde derived crosslinks. J Biomed Mater Res A 24: 1185–1201. doi: 10.1002/jbm.820240905
|
| [35] | Cornwell KG, Lei P, Andreadis ST, et al. (2007) Crosslinking of discrete self-assembled collagen threads: Effects on mechanical strength and cell-matrix interactions. J Biomed Mater Res A 80: 362–371. |
| [36] |
Yunoki S, Mori K, Suzuki T, et al. (2007) Novel elastic material from collagen for tissue engineering. J Mater Sci-Mater M 18: 1369–1375. doi: 10.1007/s10856-007-0121-6
|
| [37] |
Park SN, Park JC, Kim HO, et al. (2002) Characterization of porous collagen/hyaluronic acid scaffold modified by 1-ethyl-3-(3-dimethylaminopropyl) carbodiimide cross-linking. Biomaterials 23: 1205–1212. doi: 10.1016/S0142-9612(01)00235-6
|
| [38] |
Ma DH, Lai JY, Cheng HY, et al. (2010) Carbodiimide cross-linked amniotic membranes for cultivation of limbal epithelial cells. Biomaterials 31: 6647–6658. doi: 10.1016/j.biomaterials.2010.05.034
|
| [39] |
Piao Y, Chen B (2017) Synthesis and mechanical properties of double cross-linked gelatin-graphene oxide hydrogels. Int J Biol Macromol 101: 791–798. doi: 10.1016/j.ijbiomac.2017.03.155
|
| [40] |
Tanpichaia S, Oksmana K (2016) Cross-linked nanocomposite hydrogels based on cellulose nanocrystals and PVA: Mechanical properties and creep recovery. Compos Part A-Appl S 88: 226–233. doi: 10.1016/j.compositesa.2016.06.002
|
| [41] | DeQuach J, Lin J, Cam C, et al. (2013) Injectable skeletal muscle matrix hydrogel promotes neovascularization and muscle cell infiltration in a hindlimb ischemia model. Eur Cells Mater 23: 400–412. |
| [42] |
Massensini AR, Ghuman H, Saldin LT, et al. (2015) Concentration-dependent rheological properties of ECM hydrogel for intracerebral delivery to a stroke cavity. Acta Biomater 27: 116–130. doi: 10.1016/j.actbio.2015.08.040
|
| [43] |
Wolf MT, Daly KA, Brennan-Pierce EP, et al. (2012) A hydrogel derived from decellularized dermal extracellular matrix. Biomaterials 33: 7028–7038. doi: 10.1016/j.biomaterials.2012.06.051
|
| [44] |
Lee JS, Shin J, Park HM, et al. (2014) Liver extracellular matrix providing dual functions of two-dimensional substrate coating and three-dimensional injectable hydrogel platform for liver tissue engineering. Biomacromolecules 15: 206–218. doi: 10.1021/bm4015039
|
| [45] |
Pouliot RA, Link PA, Mikhaiel NS, et al. (2016) Development and characterization of a naturally derived lung extracellular matrix hydrogel. J Biomed Mater Res A 104: 1922–1935. doi: 10.1002/jbm.a.35726
|
| [46] | Ma L, Gao C, Mao Z, et al. (2004) Biodegradability and cell-mediated contraction of porous collagen scaffolds: the effect of lysine as a novel crosslinking bridge. J Biomed Mater Res A 71: 334–342. |
| [47] |
Wang HB, Dembo M, Wang YL (2000) Substrate flexibility regulate growth and apoptosis of normal but not transformed cells. Am J Physiol-Cell Ph 279: C1345–C1350. doi: 10.1152/ajpcell.2000.279.5.C1345
|
| [48] |
Engler AJ, Carag-Kriegr C, Johnson CP, et al. (2008) Embryonic cardiomyocytes beat best on a matrix with heart-like elasticity: scar-like rigidity inhibits beating. J Cell Sci 121: 3794–3802. doi: 10.1242/jcs.029678
|

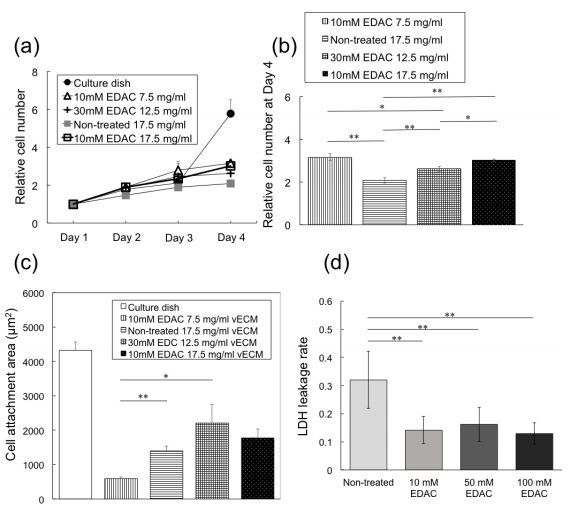
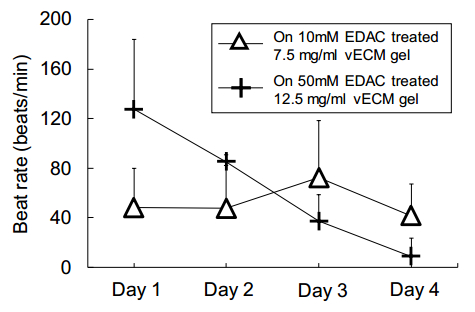
Figures(11) / Tables(1)

Kyohei Fujita, Zhonggang Feng, Daisuke Sato, Tadashi Kosawada, Takao Nakamura, Yasuyuki Shiraishi, Mitsuo Umezu. Modulation of the mechanical properties of ventricular extracellular matrix hydrogels with a carbodiimide crosslinker and investigation of their cellular compatibility[J]. AIMS Materials Science, 2018, 5(1): 54-74. doi: 10.3934/matersci.2018.1.54







 DownLoad:
DownLoad: