Citation: Gaurav Gupta, Varun Danke, Tamoor Babur, Mario Beiner. Morphology orientation of comb-like polymers with rigid backbones under the influence of shear fields[J]. AIMS Materials Science, 2017, 4(4): 970-981. doi: 10.3934/matersci.2017.4.970

| [1] |
Heeger AJ (2001) Semiconducting and metallic polymers: The fourth generation of polymeric materials. J Phys Chem B 105: 8475–8491. doi: 10.1021/jp011611w

|
| [2] |
Ong BS, Wu YL, Liu P, et al. (2004) High-performance semiconducting polythiophenes for organic thin-film transistors. J Am Chem Soc 126: 3378–3379. doi: 10.1021/ja039772w
|
| [3] |
Blom PWM, Vissenberg MCJM (2000) Charge transport in poly(p-phenylene vinylene) lightemitting diodes. Mater Sci Eng R 27: 53–94. doi: 10.1016/S0927-796X(00)00009-7
|
| [4] |
Gallot B (1996) Comb like and block liquid crystalline polymers for biological applications. Prog Polym Sci 21: 1035–1088. doi: 10.1016/S0079-6700(96)00010-X
|
| [5] |
Jianquan T, Weiqu L, Honglei W, et al. (2016) Preparation and properties of UV-curable waterborne comb-like(meth)acrylate copolymers with long fluorinated side chains. Prog Org Coat 94: 62–72. doi: 10.1016/j.porgcoat.2016.01.027
|
| [6] | Jackson WJ (1980) Liquid crystal polymers. IV. Liquid crystalline aromatic polyesters. Brit Poly J 12: 154–162. |
| [7] |
Ballauf M (1989) Stiff-chain polymers-Structure, phase-behavior, and properties. Angew Chem Int Ed Engl 28: 253–267. doi: 10.1002/anie.198902533
|
| [8] |
Shi H, Zhao Y, Dong X (2013) Frustrated crystallization and hierarchical self-assembly behavior of comb-like polymers. Chem Soc Rev 42: 2075–2099. doi: 10.1039/C2CS35350D
|
| [9] |
Beiner M, Huth H (2003) Nanophase separation and hindered glass transition in side-chain polymers. Nat Mater 2: 595–599. doi: 10.1038/nmat966
|
| [10] |
Pankaj S, Beiner M (2010) Confined dynamics and crystallization in self-assembled alkyl nanodomains. J Phys Chem B 114: 15459–15465. doi: 10.1021/jp1072999
|
| [11] |
Zheng W, Levon K, Laakso J, et al. (1994) Characterization and solid-state properties of processable N-alkylated polyanilines in the neutral state. Macromolecules 27: 7754–7768. doi: 10.1021/ma00104a034
|
| [12] | Watanabe J, Ono H, Uematsu I, et al. (1985) Thermotropic polypeptides. 2. Molecular packing and thermotropic behavior of poly(L-glutamates) with long n-alkyl side chain. Macromolecules 18: 2141–2148. |
| [13] |
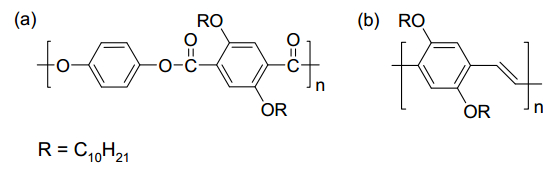
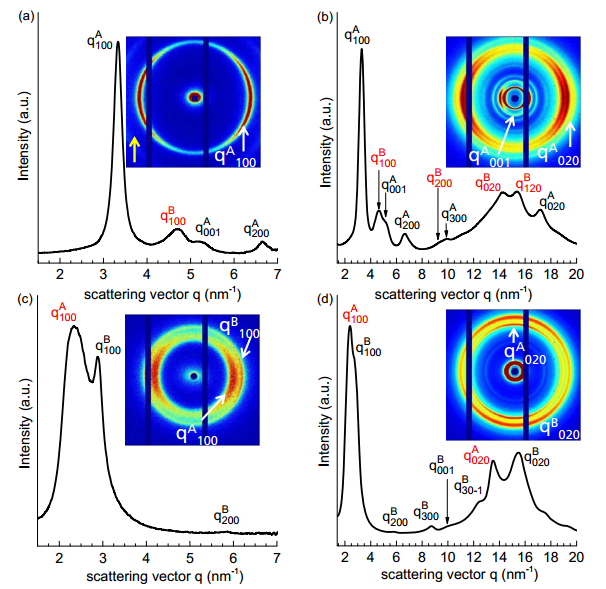
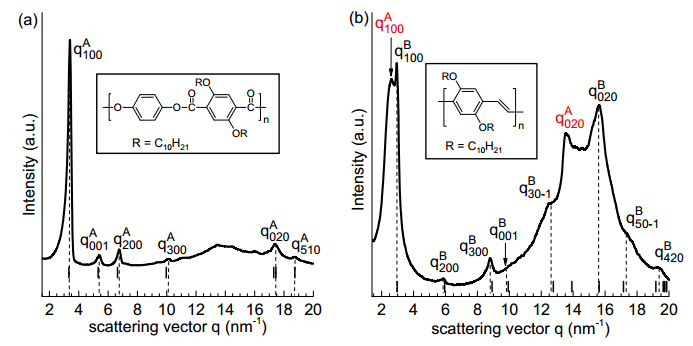
Gupta G, Danke V, Babur T, et al. (2017) Interrelations between side chain and main chain packing in different crystal modifications of alkoxylated polyesters. J Phys Chem B 121: 4583–4591. doi: 10.1021/acs.jpcb.7b00928
|
| [14] |
Sirringhaus H, Brown PJ, Friend RH, et al. (1999) Two-dimensional charge transport in selforganized, high-mobility conjugated polymers. Nature 401: 685–688. doi: 10.1038/44359
|
| [15] |
Gargi D, Kline RJ, DeLongchamp DM, et al. (2013) Charge transport in highly face-on poly(3-hexylthiophene) films. J Phys Chem C 117: 17421–17428. doi: 10.1021/jp4050644
|
| [16] |
Koynov K, Bahtiar A, Ahn T, et al. (2006) Molecular weight dependence of chain orientation and optical constants of thin films of the conjugated polymer MEH-PPV. Macromolecules 39: 8692–8698. doi: 10.1021/ma0611164
|
| [17] |
Hamaguchi M, Yoshino K (1995) Rubbing-induced molecular orientation and polarized electroluminescence in conjugated polymer. Jpn J Appl Phys 34: 712–715. doi: 10.1143/JJAP.34.712
|
| [18] |
Okuzaki H, Hirata Y, Kunugi T (1999) Mechanical properties and structure of zone-drawn poly(pphenylene vinylene) films. Polymer 40: 2625–2629. doi: 10.1016/S0032-3861(98)00511-4
|
| [19] |
Lin NT, Satyanarayana K, Chen CH, et al. (2014) Controlling the orientation of pendants in twodimensional comb-like polymers by varying stiffness of polymeric backbones. Macromolecules 47: 6166–6172. doi: 10.1021/ma5007655
|
| [20] |
Jeng U, Hsu CH, Sheu HS, et al. (2005) Morphology and charge transport in poly(2-methoxy-5-(2'-ethylhexyloxy)-1,4-phenylenevinylene) films. Macromolecules 38: 6566–6574. doi: 10.1021/ma050367u
|
| [21] |
Singh CR, Gupta G, Lohwasser R, et al. (2013) Correlation of charge transport with structural order in highly ordered melt crystallized P3HT films. J Polym Sci Part B Polym Phys 51: 943–951. doi: 10.1002/polb.23297
|
| [22] |
Brinkmann M, Contal C, Kayunkid N, et al. (2010) Highly oriented and nanotextured films of regioregular poly(3-hexylthiophene) grown by epitaxy on the nanostructured surface of an aromatic substrate. Macromolecules 43: 7604–7610. doi: 10.1021/ma101313m
|
| [23] |
Kim JS, Park Y, Lee DY, et al. (2010) Poly(3-hexylthiophene) nanorods with aligned chain orientation for organic photovoltaics. Adv Funct Mater 20: 540–545. doi: 10.1002/adfm.200901760
|
| [24] |
Fischer FSU, Tremel K, Sommer M, et al. (2012) Directed crystallization of poly(3-hexylthiophene) in micrometre channels under confinement and in electric fields. Nanoscale 4: 2138–2144. doi: 10.1039/c2nr12037b
|
| [25] |
Ebert F, Thurn-Albrecht T (2003) Controlling the orientation of semicrystalline polymers by crystallization in magnetic fields. Macromolecules 36: 8685–8694. doi: 10.1021/ma034760g
|
| [26] | Freidzon YAS, Talroze RV, Boiko NI, et al. (1988) Thermotropic liquid-crystalline polymers XXIII. Peculiarities of uniaxial orientation of comb-like liquid-crystalline polymers under mechanical stress. Liq Cryst 3: 127–132. |
| [27] |
Nagamatsu S, Takashima W, Kaneto K (2003) Backbone arrangement in friction-transferred regioregular poly(3-alkylthiophene)s. Macromolecules 36: 5252–5257. doi: 10.1021/ma025887t
|
| [28] |
Hamidi-Sakr A, Biniek L, Fall S, et al. (2016) Precise control of lamellar thickness in highly oriented regioregular poly(3-hexylthiophene) thin films prepared by high-temperature rubbing: Correlations with optical properties and charge transport. Adv Funct Mater 26: 408–420. doi: 10.1002/adfm.201504096
|
| [29] |
Rim YS, Bae SH, Chen HJ, et al. (2016) Recent progress in materials and devices toward Printable and flexible sensors. Adv Mater 28: 4415–4440. doi: 10.1002/adma.201505118
|
| [30] |
Reinspach JA, Diao Y, Giri G, et al. (2016) Tuning the morphology of solution-sheared P3HT:PCBM films. ACS Appl Mater Interfaces 8: 1742–1751. doi: 10.1021/acsami.5b09349
|
| [31] | Rauwendaal C (2001) Polymer Extrusion, Carl Hanser Verlag Munich. |
| [32] | Damman SB, Vroege GJ (1993) Liquid-crystalline main-chain polymers with a poly(p-phenylene terephthalate) backbone. X-ray-diffraction of the polyester with dodecyloxy side-chains. Polymer 34: 2732–2739. |
| [33] |
Babur T, Balko J, Budde H, et al. (2014) Confined relaxation dynamics in long range ordered polyesters with comb-like architecture. Polymer 55: 6844–6852. doi: 10.1016/j.polymer.2014.10.062
|
| [34] | Babur T (2017) Structure and relaxation dynamics of comb-like polymers with rigid backbone [PhD Dissertation]. Martin Luther University Halle-Wittenberg. |
| [35] |
Babur T, Gupta G, Beiner M (2016) About different packing states of alkyl groups in comb-like polymers with rigid backbones. Soft Matter 12: 8093–8097. doi: 10.1039/C6SM01812B
|
| [36] |
Nagamatsu S, Misaki M, Chikamatsu M, et al. (2007) Crystal structure of friction transferred poly(2,5-dioctyloxy-1,4-phenylenevinylene). J Phys Chem B 111: 4349–4354. doi: 10.1021/jp067555m
|
| [37] |
Balsara NP, Dai HJ (1996) A transition from shear-induced order to shear-induced disorder in block copolymers. J Chem Phys 105: 2942–2945. doi: 10.1063/1.472162
|
| [38] | Vigild ME, Chu C, SugiyamaM(2001) Influence of shear on the alignment of a lamellae-forming pentablock Copolymer. Macromolecules 34: 951–964. |
| [39] | Damman SB, Mercx FPM (1993) About di erent packing states of alkyl groups in comb-like polymers with rigid backbones. J Polym Sci Part B Polym Phys 31: 1759–1767. |
| [40] | Falk U, Westermark B, Boeffel C, et al. (1987) NMR of stiff macromolecules with flexible side chains. Mol Cryst Liq Cryst 153: 199–206. |
Figures(6) / Tables(1)

Gaurav Gupta, Varun Danke, Tamoor Babur, Mario Beiner. Morphology orientation of comb-like polymers with rigid backbones under the influence of shear fields[J]. AIMS Materials Science, 2017, 4(4): 970-981. doi: 10.3934/matersci.2017.4.970







 DownLoad:
DownLoad: