Citation: Shota Tobaru, Ryuto Shigenobu, Foday Conteh, Naomitsu Urasaki, Abdul Motin Howlader, Tomonobu Senjyu, Toshihisa Funabashi. Optimal operation method coping with uncertainty in multi-area small power systems[J]. AIMS Energy, 2017, 5(4): 718-734. doi: 10.3934/energy.2017.4.718

| [1] |
Amenedo JLR, Arnalte S, Burgos JC, et al. (2002) Automatic generation control of a wind farm with variable speed wind turbines. IEEE T Energy Conver 17: 279–284. doi: 10.1109/TEC.2002.1009481

|
| [2] | Senjyu T, Hayashi D, Sakamoto Y, et al. (2005) Generating power leveling of renewable energy for small power system in isolated island. IEE Japan 125: 1209–1215. |
| [3] | Senjyu T, Tokudome M, Yona A, et al. (2009) A Frequency Control Approach by Decentralized Controllable Loads in Small Power Systems. IEE Japan 129: 1074–1080. |
| [4] | Senjyu T, Kikunaga T, Tokudome M, et al. (2009) Coordinate Control ofWind Turbine and Battery in Wind Turbine Generator System. IEE Japan 129: 653–660. |
| [5] | Kurohane K, Senjyu T, Yona A, et al. (2010) A High Quality Power Supply System with DC Smart Grid. 2010 IEEE PES Transmission and Distribution Conference and Exposition. |
| [6] | Ogimi K, Kamiyama S, Palmer M, et al. (2013) Optimal OperationPlanning ofWind Farm Installed BESS Using Wind Power Forecast Data of Wind Turbine Generators Considering Forecast Error. Int J Emerg Electric Power Syst 14: 207–218. |
| [7] | Bracale A, Carpinelli G, Fazio AD, et al. (2015) Advanced, Cost-Based Indices for Forecasting the Generation of Photovoltaic Power. Int J Emerg Electric Power Syst 16: 195–206. |
| [8] | Shimoji T, Tahara H, Matayoshi H, et al. (2015) Comparison and Validation of Operational Cost in Smart Houses with the Introduction of a Heat Pump or a Gas Engine. Int J Emerg Electric Power Syst 16: 59–74. |
| [9] | MengyanW, HigaW, Yona A, et al. (2012) Optimal Operation of Power Systems with Power Players. 2012 International Conference on Renewable Energy Research and Applications (ICRERA). |
| [10] | Higa S, Mengyan W, Yona A, et al. (2013) Optimal Operation Method Considering Uncertainly of Renewable Energy and Load Demand in Micro-grids. The 5th International Conference on Advanced Power System Automation and Protection. |
| [11] | Higa S, Howlader AM, Shiroma Y, et al. (2014) Optimal Operation Method Considering Replanning and Uncertainly in Power Systems. The International Conference on Electrical Engineering 2014. |
| [12] |
Zheng Y, Dong ZY, Luo FJ, et al. (2014) Optimal Allocation of Energy Storage System for Risk Mitigation of DISCOs With High Renewable Penetrations. IEEE T Power Syst 29: 212–220. doi: 10.1109/TPWRS.2013.2278850
|
| [13] |
Zhang L, Li Y (2013) Optimal Energy Management of Wind-Battery Hybrid Power System With Two-Scale Dynamic Programming. IEEE T Sust Energy 4: 765–773. doi: 10.1109/TSTE.2013.2246875
|
| [14] |
Ahn SJ, Nam SR, Choi JH, et al. (2013) Power Scheduling of Distributed Generators Economic and Stable Operation of a Microtrid. IEEE T Smart Grid 4: 398–405. doi: 10.1109/TSG.2012.2233773
|
| [15] |
Zhao B, Zhang X, Chen J, et al. (2013) Operation Optimization of Standalone Microgrids Considering Lifetime Characteristics of Battery Energy StorageSystem. IEEE T Sust Energy 4: 934–943. doi: 10.1109/TSTE.2013.2248400
|
| [16] | He D, Tan Z, Harley RG, et al. (2012) Chance Constrained Unit Commitment With Wind Generation and Superconducting Magnetic Energy Storages. 2012 IEEE Power and Energy Society General Meeting. |
| [17] | Reddy SS, Abhyankar AR, Bijwe PR, et al. (2014) Co-optimization of Energy and Demand-Side Reserves in Day-Ahead Electricity Markets. Int J Emerg Electric Power Syst 15: 77–91. |
| [18] | Nguyen MY, Nguyen DM (2015) A Generalized Formulation of Demand Response under Market Environments. Int J Emerg Electric Power Syst 16: 217–224. |
| [19] |
Aalami HA, Moghaddam MP, Yousefi GR, et al. (2010) Demand response modeling considering Interruptible/Curtailable loads and capacity market programs. Appl Energy 87: 243–250. doi: 10.1016/j.apenergy.2009.05.041
|
| [20] | Andebili MR, Shen H (2017) Energy Management of End Users Modeling their Reaction from a GENCOs Point of View. 2017 International Conference on Computing, Networking and Communications (ICNC). |
| [21] |
Andebili MR (2016) Modeling nonlinear incentive-based and price-based demand response programs and implementing on real power markets. Electr Pow Syst Res 132: 115–124. doi: 10.1016/j.epsr.2015.11.006
|
| [22] | Andebili MR, Shen H (2017) Price-Controlled Energy Management of Smart Homes for Maximizing Profit of a GENCO. IEEE T Syst Man Cy A. |
| [23] |
Andebili MR (2015) Risk-Cost Based Generation Scheduling Smartly Mixed with Reliability and Market-Driven Demand Response Measures. Int T Electr Energy Syst 25: 994–1007. doi: 10.1002/etep.1884
|
| [24] | AndebiliMR(2016) Nonlinear demand response programs for residential customers with nonlinear behavioral models. Energ Buildings 119: 352–362. |
| [25] |
Andebili MR (2013) Investigating effects of responsive loads models on unit commitment collaborated with demand-side resources. IET Gener Transm D 7: 420–430. doi: 10.1049/iet-gtd.2012.0552
|
| [26] | Japan Electric Power Exchange, JEPX, 2017. Available from: http://www.jepx.org/index.html. |
| [27] |
Nguyen MY, Yoon YT (2014) Optimal Scheduling and Operation of Battery/Wind Generation System in Response to Real-time Market Prices. IEE T Electr Electr Eng 9: 129–135. doi: 10.1002/tee.21947
|
| [28] |
Havel P (2014) Utilization of Real-Time Balancing Market for Transmission System Control Under Uncertainty. IEEE T Power Syst 29: 450–457. doi: 10.1109/TPWRS.2012.2228676
|
| [29] | Kiviluoma J, O'Malley M, Tuohy A, et al. (2011) Impact ofWind Power on the Unit Commitment, Operating Reserves, and Market Design. 2011 IEEE Power and Energy Society General Meeting. |
| [30] | Vlachos AG, Biskas PN, et al. (2011) Balancing Supply and Demand Under Mixed Pricing Rules in Multi-Area Electricity Markets. IEEE T Power Syst 26: 1444–1453. |
| [31] | Ma J, Silva V, Belhomme R, et al. (2013) Evaluating and Planning Flexibility in Sustainable Power Systems. 2013 IEEE Power and Energy Society General Meeting (PES). |
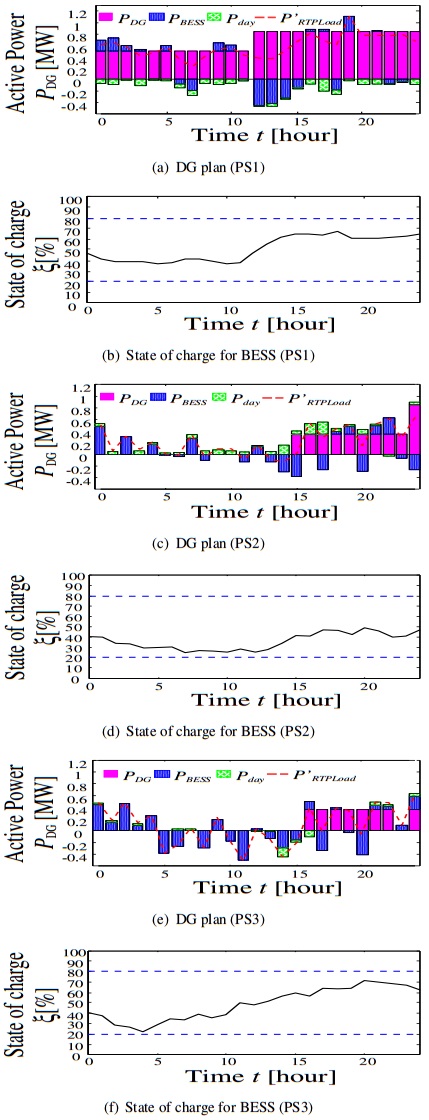
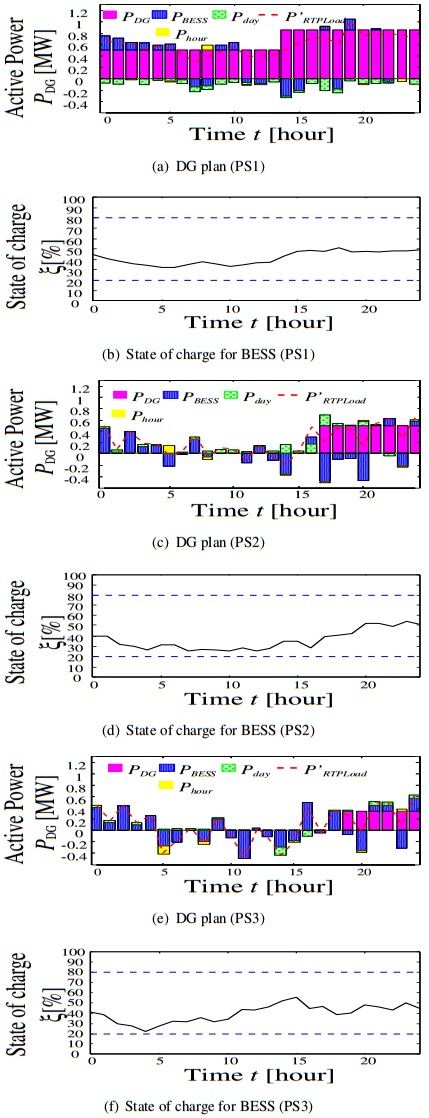
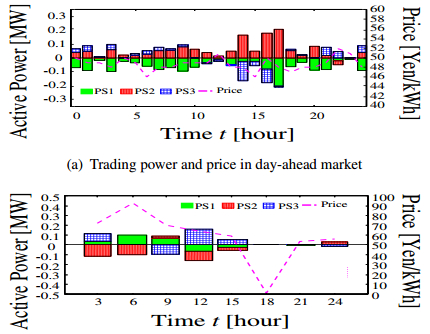
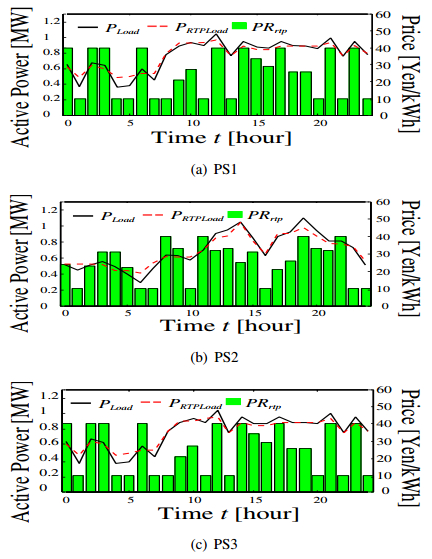
Figures(12) / Tables(2)

Shota Tobaru, Ryuto Shigenobu, Foday Conteh, Naomitsu Urasaki, Abdul Motin Howlader, Tomonobu Senjyu, Toshihisa Funabashi. Optimal operation method coping with uncertainty in multi-area small power systems[J]. AIMS Energy, 2017, 5(4): 718-734. doi: 10.3934/energy.2017.4.718







 DownLoad:
DownLoad: