Citation: Grigorios L. Kyriakopoulos, Garyfallos Arabatzis, Miltiadis Chalikias. Renewables exploitation for energy production and biomass use for electricity generation. A multi-parametric literature-based review[J]. AIMS Energy, 2016, 4(5): 762-803. doi: 10.3934/energy.2016.5.762

| [1] | Batel S, Devine-Wright P (2015) Towards a better understanding of people’s responses to renewable energy technologies: Insights from Social Representations Theory. Public Und Sci 24: 311-325. |
| [2] | Devine-Wright P (2010) Place attachment and the social acceptance of renewable energy technologies. Chapter 17 In: Psychological Approaches to Sustainability: Current Trends in Theory, Research and Applications. Edited by Victor Corral-Verdugo and Cirilo H. Garcia-Cadena. Nova Science Publishers Eds., 317-336. |
| [3] |
Mallett A (2007) Social acceptance of renewable energy innovations: The role of technology cooperation in urban Mexico. Energ Policy 35: 2790-2798. doi: 10.1016/j.enpol.2006.12.008

|
| [4] | Groba F, Cao J (2014) Chinese Renewable Energy Technology Exports: The Role of Policy, Innovation and Markets. Environ Resour Econ 60: 243-283. |
| [5] |
Sung B, Song WY (2014) How government policies affect the export dynamics of renewable energy technologies: A subsectoral analysis. Energy 69: 843-859. doi: 10.1016/j.energy.2014.03.082
|
| [6] |
Zhang X, Chang S, Eric M (2012) Renewable energy in China: An integrated technology and policy perspective. Energ Policy 51: 1-6. doi: 10.1016/j.enpol.2012.09.071
|
| [7] | Negro SO, Hekkert MP, Smits REHM (2008) Stimulating renewable energy technologies by innovation policy. Sci Public Policy 35: 403-416. |
| [8] | Jacobsson S, Lauber V (2006) The politics and policy of energy system transformation - Explaining the German diffusion of renewable energy technology. Energ Policy 34: 256-276. |
| [9] |
Tsoutsos TD, Stamboulis YA (2005) The sustainable diffusion of renewable energy technologies as an example of an innovation-focused policy. Technovation 25: 753-761. doi: 10.1016/j.technovation.2003.12.003
|
| [10] | Buckley JC, Schwarz PM (2003) Renewable energy from gasification of manure: An innovative technology in search of fertile policy. Environ Monit Assess 84: 111-127. |
| [11] | Hervás Soriano F, Mulatero F (2011) EU Research and Innovation (R&I) in renewable energies: The role of the Strategic Energy Technology Plan (SET-Plan). Energ Policy 39: 3582-3590. |
| [12] |
Shen YC, Lin GTR, Li KP, et al. (2010) An assessment of exploiting renewable energy sources with concerns of policy and technology. Energ Policy 38: 4604-4616. doi: 10.1016/j.enpol.2010.04.016
|
| [13] |
Van Alphen K, Kunz HS, Hekkert MP (2008) Policy measures to promote the widespread utilization of renewable energy technologies for electricity generation in the Maldives. Renew Sust Energ Rev 12: 1959-1973. doi: 10.1016/j.rser.2007.04.009
|
| [14] | Gan J, Smith CT (2006) A comparative analysis of woody biomass and coal for electricity generation under various CO2 emission reductions and taxes. Biomass Bioenerg 30: 296-303. |
| [15] |
Sung B, Song WY (2013) Causality between public policies and exports of renewable energy technologies. Energ Policy 55: 95-104. doi: 10.1016/j.enpol.2012.10.063
|
| [16] | Garces E, Daim T (2013) An assessment of the impacts of government energy policy on energy technology, innovation, and security: The case of renewable technologies in the US electricity sector. Lect Notes Energ 16: 257-276. |
| [17] |
Anderson D (1997) Renewable energy technology and policy for development. Annu Rev Energ Env 22: 187-215. doi: 10.1146/annurev.energy.22.1.187
|
| [18] | Nisar A, Ruiz F, Palacios M (2013) Organisational learning, strategic rigidity and technology adoption: Implications for electric utilities and renewable energy firms. Renew Sust Energ Rev 22: 438-445. |
| [19] | García VG, Bartolomé MM (2010) Rural electrification systems based on renewable energy: The social dimensions of an innovative technology. Technol Soc 32: 303-311. |
| [20] |
Tsai WT (2005) Current status and development policies on renewable energy technology research in Taiwan. Renew Sust Energ Rev 9: 237-253. doi: 10.1016/j.rser.2004.03.003
|
| [21] | Abidin NZZ, Ibrahim JB (2015) Embracing renewables - Overcoming integration challenges from Malaysia's utility perspective. 2015 IEEE Eindhoven PowerTech, PowerTech, 2015: art. no. 7232373. |
| [22] | Loiter JM, Norberg-Bohm V (1999) Technology policy and renewable energy: Public roles in the development of new energy technologies. Energ Policy 27: 85-97. |
| [23] | Franco A, Salza P (2011) Perspectives for the long-term penetration of new renewables in complex energy systems: The Italian scenario. Environ Dev Sustainability 13: 309-330. |
| [24] | Amorim F, Martins MVM, Pereira Da Silva P (2010) A new perspective to account for renewables impacts in Portugal. 7th International Conference on the European Energy Market 2010, EEM 2010: art. no. 5558695. |
| [25] |
Suwa A, Jupesta J (2012) Policy innovation for technology diffusion: A case-study of Japanese renewable energy public support programs. Sustainability Sci 7: 185-197. doi: 10.1007/s11625-012-0175-3
|
| [26] |
Vassilev SV, Vassileva CG, Vassilev VS (2015) Advantages and disadvantages of composition and properties of biomass in comparison with coal: An overview. Fuel 158: 330-350. doi: 10.1016/j.fuel.2015.05.050
|
| [27] | Zandler H, Brenning A, Samimi C (2015) Potential of space-borne hyperspectral data for biomass quantification in an arid environment: Advantages and limitations. Remote Sens 7: 4565-4580. |
| [28] | Costa C, Rotsios K, Moshonas G, et al. (2014) Use of multivariate approaches in biomass energy plantation harvesting: Logistics advantages. Agr Eng Int: CIGR Journal May 2014: 71-80. |
| [29] | Bick S (2013) Skidding options for biomass operations: From big to small, each alternative has advantages and drawbacks. Northern Logger Timber Processor 62: 30-35. |
| [30] | Vassilev SV, Baxter D, Andersen LK, et al. (2013) An overview of the composition and application of biomass ash.: Part 2. Potential utilisation, technological and ecological advantages and challenges. Fuel 105: 19-39. |
| [31] | Talluri S, Subramanian MR, Christopher LP (2012) Advantages of thermophilic hydrogen production from lignocellulosic biomass. 2012 AIChE Annual Meeting Conference Proceedings: 1p. |
| [32] | Lanning CJ, Fridley JL (2012) Advances in woody biomass drying by taking advantage of surface properties. American Society of Agricultural and Biological Engineers Annual International Meeting 2012, ASABE 2012, 1: 715-723. |
| [33] |
Santos CA, Ferreira ME, Lopes Da Silva T, et al. (2011) A symbiotic gas exchange between bioreactors enhances microalgal biomass and lipid productivities: Taking advantage of complementary nutritional modes. J Ind Microbiol Biotechn 38: 909-917. doi: 10.1007/s10295-010-0860-0
|
| [34] | Hotchkiss R, Matts D, Riley G (2003) Co-combustion of Biomass with Coal - The Advantages and Disadvantages Compared to Purpose-built Biomass to Energy Plants. VGB PowerTech 83: 80-86. |
| [35] |
Fransen B, De Kroon H (2001) Long-term disadvantages of selective root placement: Root proliferation and shoot biomass of two perennial grass species in a 2-year experiment. J Ecol 89: 711-722. doi: 10.1046/j.0022-0477.2001.00589.x
|
| [36] |
Nishiguchi S, Tabata T (2016) Assessment of social, economic, and environmental aspects of woody biomass energy utilization: Direct burning and wood pellets. Renew Sust Energ Rev 57: 1279-1286. doi: 10.1016/j.rser.2015.12.213
|
| [37] |
Miret C, Chazara P, Montastruc L, et al. (2016) Design of bioethanol green supply chain: Comparison between first and second generation biomass concerning economic, environmental and social criteria. Comput Chem Eng 85: 16-35. doi: 10.1016/j.compchemeng.2015.10.008
|
| [38] |
Singh J (2015) Overview of electric power potential of surplus agricultural biomass from economic, social, environmental and technical perspective - A case study of Punjab. Renew Sust Energ Rev 42: 286-297. doi: 10.1016/j.rser.2014.10.015
|
| [39] |
Cambero C, Sowlati T (2014) Assessment and optimization of forest biomass supply chains from economic, social and environmental perspectives - A review of literature. Renew Sust Energ Rev 36: 62-73. doi: 10.1016/j.rser.2014.04.041
|
| [40] |
Aylott MJ, Casella E, Farrall K, et al. (2010) Estimating the supply of biomass from short-rotation coppice in England, given social, economic and environmental constraints to land availability. Biofuels 1: 719-727. doi: 10.4155/bfs.10.30
|
| [41] |
Haughton AJ, Bond AJ, Lovett AA, et al. (2009) A novel, integrated approach to assessing social, economic and environmental implications of changing rural land-use: A case study of perennial biomass crops. J Appl Ecol 46: 315-322. doi: 10.1111/j.1365-2664.2009.01623.x
|
| [42] |
Fischer SL, Koshland CP, Young JA (2005) Social, economic, and environmental impacts assessment of a village-scale modern biomass energy project in Jilin province, China: local outcomes and lessons learned. Energ Sust Dev 9: 50-59. doi: 10.1016/S0973-0826(08)60499-8
|
| [43] |
Felker P (1984) Economic, environmental, and social advantages of intensively managed short rotation mesquite (Prosopis spp) biomass energy farms. Biomass 5: 65-77. doi: 10.1016/0144-4565(84)90070-2
|
| [44] |
Yilmaz Balaman T, Selim H (2015) A decision model for cost effective design of biomass based green energy supply chains. Bioresource Technol 191: 97-109. doi: 10.1016/j.biortech.2015.04.078
|
| [45] | Dillibabu V, Natarajan E (2015) Green energy from biomass gasification. J Chem Pharm Sci 7: 182-185. |
| [46] |
Hamzeh Y, Ashori A, Mirzaei B, et al. (2011) Current and potential capabilities of biomass for green energy in Iran. Renew Sust Energ Rev 15: 4934-4938. doi: 10.1016/j.rser.2011.07.060
|
| [47] |
Bhutto AW, Bazmi AA, Zahedi G (2011) Greener energy: Issues and challenges for Pakistan - Biomass energy prospective. Renew Sust Energ Rev 15: 3207-3219. doi: 10.1016/j.rser.2011.04.015
|
| [48] | Demirbas T, Demirbas AH (2010) Bioenergy, green energy. biomass and biofuels. Energ Sources 32: 1067-1075. |
| [49] | Kaltschmitt M, Thrän D (2009) Biomass-based green energy generation. Chapter 7 In: Sustainable Solutions for Modern Economies. Edited by Martin Kaltschmitt and Daniela Thrän. Royal Society of Chemistry (RSC) Green Chemistry Eds., 86-124. |
| [50] | Jenner M (2008) Biomass energy outlook: How green is your carbon? BioCycle 49: 41. |
| [51] |
Alfonsin V, Suarez A, Urrejola S, Miguez J, Sanchez A (2015) Integration of several renewable energies for internal combustion engine substitution in a commercial sailboat. Int J Hydrogen Energ 40: 6689-6701. doi: 10.1016/j.ijhydene.2015.02.113
|
| [52] |
Iniyan S, Suganthi L, Samuel AA (2006) Energy models for commercial energy prediction and substitution of renewable energy sources. Energ Policy 34: 2640-2653. doi: 10.1016/j.enpol.2004.11.017
|
| [53] | Lombard A, Ferreira SLA (2015) The spatial distribution of renewable energy infrastructure in three particular provinces of South Africa. Bull Geogr 30: 71-85. |
| [54] |
Palmas C, Siewert A, von Haaren C (2015) Exploring the decision-space for renewable energy generation to enhance spatial efficiency. Environ Impact Assess Rev 52: 9-17. doi: 10.1016/j.eiar.2014.06.005
|
| [55] |
Young M (2015) Building the blue economy: The role of marine spatial planning in facilitating offshore renewable energy development. Int J Marine Coastal Law 30: 148-173. doi: 10.1163/15718085-12341339
|
| [56] | Güngör-Demirci G (2015) Spatial analysis of renewable energy potential and use in Turkey. J Renew Sust Energ 2015, 7: art. no. 013126. |
| [57] |
Campbell MS, Stehfest KM, Votier SC, Hall-Spencer JM (2014) Mapping fisheries for marine spatial planning: Gear-specific vessel monitoring system (VMS), marine conservation and offshore renewable energy. Marine Policy 45: 293-300. doi: 10.1016/j.marpol.2013.09.015
|
| [58] |
Karakostas S, Economou D (2014) Enhanced multi-objective optimization algorithm for renewable energy sources: Optimal spatial development of wind farms. Int J Geogr Inform Sci 28: 83-103. doi: 10.1080/13658816.2013.820829
|
| [59] |
Wang Q, M’Ikiugu MM, Kinoshita I (2014) A GIS-based approach in support of spatial planning for renewable energy: A case study of Fukushima, Japan. Sustainability 6: 2087-2117. doi: 10.3390/su6042087
|
| [60] |
Behringer S, Upmann T (2014) Optimal harvesting of a spatial renewable resource. J Econ Dyn Control 42: 105-120. doi: 10.1016/j.jedc.2014.03.008
|
| [61] |
Davies IM, Watret R, Gubbins M (2014) Spatial planning for sustainable marine renewable energy developments in Scotland. Ocean Coastal Manage 99: 72-81. doi: 10.1016/j.ocecoaman.2014.05.013
|
| [62] |
Yang C, Ogden JM (2013) Renewable and low carbon hydrogen for California-Modeling the long term evolution of fuel infrastructure using a quasi-spatial TIMES model. Int J Hydrogen Energ 38: 4250-4265. doi: 10.1016/j.ijhydene.2013.01.195
|
| [63] | Baltas AE, Dervos AN (2012) Special framework for the spatial planning & the sustainable development of renewable energy sources. Renew Energ 48: 358-363. |
| [64] |
Haller M, Ludig S, Bauer N (2012) Decarbonization scenarios for the EU and MENA power system: Considering spatial distribution and short term dynamics of renewable generation. Energ Policy 47: 282-290. doi: 10.1016/j.enpol.2012.04.069
|
| [65] | Katsaprakakis DA, Christakis DG (2016) The exploitation of electricity production projects from Renewable Energy Sources for the social and economic development of remote communities. the case of Greece: An example to avoid. Renew Sust Energ Rev 54:341-349. |
| [66] | Kyriakopoulos GL, Arabatzis G (2016) Electrical energy storage systems in electricity generation: Energy policies, innovative technologies, and regulatory regimes. Renew Sust Energ Rev 56:1044-1067. |
| [67] |
Zografidou E, Petridis K, Arabatzis G, et al. (2016) Optimal design of the renewable energy map of Greece using weighted goal-programming and data envelopment analysis. Comput Oper Res 66: 313-326. doi: 10.1016/j.cor.2015.03.012
|
| [68] |
Ntona E, Arabatzis G, Kyriakopoulos GL (2015) Energy saving: Views and attitudes of students in secondary education. Renew Sust Energ Rev 46: 1-15. doi: 10.1016/j.rser.2015.02.033
|
| [69] |
Arabatzis G, Petridis K, Galatsidas S, et al. (2013) A demand scenario based fuelwood supply chain: A conceptual model. Renew Sust Energ Rev 25: 687-697. doi: 10.1016/j.rser.2013.05.030
|
| [70] |
Arabatzis G, Malesios Ch (2013) Pro-Environmental attitudes of users and not users of fuelwood in a rural area of Greece. Renew Sust Energ Rev 22: 621-630. doi: 10.1016/j.rser.2013.02.026
|
| [71] | Balaras CA, Dascalaki EG, Droutsa P, et al. (2013) Hellenic renewable energy policies and energy performance of residential buildings using solar collectors for domestic hot water production in Greece. J Renew Sust Energ 5: art. no. 041813. |
| [72] | Kyriakopoulos G, Chalikias M (2013) The Investigation of Woodfuels’ Involvement in Green Energy Supply Schemes at Northern Greece: The Model Case of the Thrace Prefecture. Procedia Technol 8: 445-452. |
| [73] | Menegaki AN, Gurluk S (2013) Greece & Turkey; assessment and comparison of their renewable energy performance. Int J Energ Econ Policy 3: 367-383. |
| [74] | Metaxas A, Tsinisizelis M (2013) The development of renewable energy governance in Greece. Examples of a failed (?) policy. Lecture Notes Energy 23: 155-168. |
| [75] |
Mondol JD, Koumpetsos N (2013) Overview of challenges, prospects, environmental impacts and policies for renewable energy and sustainable development in Greece. Renew Sust Energ Rev 23: 431-442. doi: 10.1016/j.rser.2013.01.041
|
| [76] |
Zafirakis D, Chalvatzis K, Kaldellis JK (2013) Socially just support mechanisms for the promotion of renewable energy sources in Greece. Renew Sust Energ Rev 21: 478-493. doi: 10.1016/j.rser.2012.12.030
|
| [77] |
Arabatzis G, Kitikidou K, Tampakis S, et al. (2012) The fuelwood consumption in a rural area of Greece. Renew Sust Energ Rev 16: 6489-6496. doi: 10.1016/j.rser.2012.07.010
|
| [78] | Chalikias Μ, Kyriakopoulos G, Goulionis J, et al. (2012). Investigation of the parameters affecting fuelwoods’ consumption in the Southern Greece region. J Food Agr Environ 10: 885-889. |
| [79] |
Kaldellis JK, Kapsali M, Katsanou E (2012) Renewable energy applications in Greece-What is the public attitude? Energ Policy 42: 37-48. doi: 10.1016/j.enpol.2011.11.017
|
| [80] | Markatou M (2012) Renewable energy technologies in greece: A patent based approach. Int J Renew Energ Res 2: 718-722. |
| [81] | Mourmouris JC, Potolias C, Jacob FG (2012) Evaluation of renewable energy sources exploitation at remote regions, using computing model and multi-criteria analysis: A case-study in samothrace, Greece. Int J Renew Energ Res 2: 307-316. |
| [82] |
Tegou LI, Polatidis H, Haralambopoulos DA (2012) A multi-criteria framework for an isolated electricity system design with renewable energy sources in the context of distributed generation: The case study of Lesvos Island, Greece. Int J Green Energ 9: 256-279. doi: 10.1080/15435075.2011.621484
|
| [83] | Kolovos K, Kyriakopoulos G, Chalikias M (2011). Co-evaluation of basic woodfuel types used as alternative heating sources to existing energy network. J Environ Protect Ecol 12: 733-742. |
| [84] |
Tourkolias C, Mirasgedis S (2011) Quantification and monetization of employment benefits associated with renewable energy technologies in Greece. Renew Sust Energ Rev 15: 2876-2886. doi: 10.1016/j.rser.2011.02.027
|
| [85] |
Boemi SN, Papadopoulos AM, Karagiannidis A, et al. (2010) Barriers on the propagation of renewable energy sources and sustainable solid waste management practices in Greece. Waste Manage Res 28: 967-976. doi: 10.1177/0734242X10375867
|
| [86] | Chalikias M, Kyriakopoulos G, Kolovos K (2010). Environmental sustainability and financial feasibility evaluation of woodfuel biomass used for a potential replacement of conventional space heating sources. Part I: A Greek Case Study. Oper Res 10: 43-56. |
| [87] | Kyriakopoulos G (2010) European and international policy interventions of implementing the use of woodfuels in bioenergy sector. A trend analysis and a specific woodfuels’ energy application. Int J Knowl Learn 6: 43-54. |
| [88] | Kyriakopoulos G, Kolovos K, Chalikias M (2010) Environmental sustainability and financial feasibility evaluation of woodfuel biomass used for a potential replacement of conventional space heating sources. Part II: A Combined Greek and the nearby Balkan Countries Case Study. Oper Res 10: 57-69. |
| [89] |
Kyriakopoulos G (2009) Biomass utilization for energy infrastructure and applications. Int J Soc Humanistic Comput 1: 163-174. doi: 10.1504/IJSHC.2009.031005
|
| [90] | Tsiblostefanakis EC, Mantouka KA (2009) Economy efficiency for renewable energy sources in Greece. WSEAS T Bus Econ 6: 42-51. |
| [91] |
Zafeiriou E, Arabatzis G, Koutroumanidis T (2011) The fuelwood market in Greece: An empirical approach. Renew Sust Energ Rev 15: 3008-3018. doi: 10.1016/j.rser.2011.03.019
|
| [92] |
Xydis G, Koroneos C (2009) Alternative scenarios of the utilisation of renewable energy sources in small prefectures: A case study in lasithi prefecture, Greece. Int J Glob Energ Issues 31: 61-87. doi: 10.1504/IJGEI.2009.021543
|
| [93] | Angelis-Dimakis A, Trogadas P, Arampatzis G, et al. (2008) Cost Effectiveness Analysis for Renewable Energy Sources integration in the island of Lemnos, Greece. Proceedings iEMSs 4th Biennial Meeting - International Congress on Environmental Modelling and Software: Integrating Sciences and Information Technology for Environmental Assessment and Decision Making - iEMSs 2008: 1215-1222. |
| [94] | Lazarou S, Noou K, Siassiakos K, et al. (2008) The impact of renewable energy sources penetration in achieving the energy and environmental policy goals in Greece. WSEAS T Environ Dev 4: 1161-1170. |
| [95] |
Tsoutsos T, Papadopoulou E, Katsiri A, et al. (2008) Supporting schemes for renewable energy sources and their impact on reducing the emissions of greenhouse gases in Greece. Renew Sust Energ Rev 12: 1767-1788. doi: 10.1016/j.rser.2007.04.002
|
| [96] | Mirasgedis S, Sarafidis Y, Georgopoulou E, et al. (2002) The role of renewable energy sources within the framework of the Kyoto Protocol: The case of Greece. Renew Sust Energ Rev 6: 249-272. |
| [97] | Jäger-Waldau A, Szabó M, Monforti-Ferrario F, et al. (2011) Renewable energy snapshots 2011. JRC Scientific and Technical Reports of European Commission – Joint Research Centre Institute for Energy and Transport. Available from: http://iet.jrc.ec.europa.eu/remea/renewable-energy-snapshots-2011. |
| [98] | Schenk T, Stokes LC (2013) The power of collaboration: Engaging all parties in renewable energy infrastructure development. IEEE Power Energ Mag 11: 56-65. |
| [99] | Sasaki H, Kajikawa Y, Fujisue K, et al. (2010) Detecting the valley of international academic collaboration in renewable energy. IEEM2010 - IEEE International Conference on Industrial Engineering and Engineering Management 2010; art. no. 5674434: 99-103. |
| [100] |
Chineke TC, Ezike FM (2010) Political will and collaboration for electric power reform through renewable energy in Africa. Energ Policy 38: 678-684. doi: 10.1016/j.enpol.2009.10.004
|
| [101] |
Scarlat N, Dallemand JF, Monforti-Ferrario F, et al. (2015) Renewable Energ Policy framework and bioenergy contribution in the European Union - An overview from National Renewable Energy Action Plans and Progress Reports. Renew Sust Energ Rev 51: 969-985. doi: 10.1016/j.rser.2015.06.062
|
| [102] | Scarlat N, Dallemand JF, Banja M (2013) Possible impact of 2020 bioenergy targets on European Union land use. A scenario-based assessment from national renewable energy action plans proposals. Renew Sust Energ Rev 18: 595-606. |
| [103] | Pozeb V, Goričanec D, Krope J (2011) European Energ Policy - Directives and action plans. Recent Researches in Geography, Geology, Energy, Environment and Biomedicine. Proceedings of the 4th WSEAS International Conference on EMESEG 2011:107-110. |
| [104] |
McKillop A (2012) Europe’s climate energy package must be reformed or abandoned. Energ Environ 23: 837-848. doi: 10.1260/0958-305X.23.5.837
|
| [105] | Kulovesi K, Morgera E, Muñoz M (2011) Environmental integration and multi-faceted international dimensions of EU law: Unpacking the EU's 2009 climate and energy package. Common Market Law Rev 48: 829-891. |
| [106] | Uusi-Rauva C, Tienari J (2010) On the relative nature of adequate measures: Media representations of the EU energy and climate package. Glob Environ Change 20: 492-501. |
| [107] |
Uusi-Rauva C (2010) The EU energy and climate package: A showcase for European environmental leadership? Environ Policy Governance 20: 73-88. doi: 10.1002/eet.535
|
| [108] | Vielle M, Moulinier JM, Bernard A (2009) Evaluation of the energy-climate package with the help of gemini-e3 model. Revue de l'Energie 589: 153-173. |
| [109] |
Connolly D, Lund H, Mathiesen BV, et al. (2014) Heat roadmap Europe: Combining district heating with heat savings to decarbonise the EU energy system. Energ Policy 65: 475-489. doi: 10.1016/j.enpol.2013.10.035
|
| [110] |
Alonso PM, Hewitt R, Pacheco JD, et al. (2016) Losing the roadmap: Renewable energy paralysis in Spain and its implications for the EU low carbon economy. Renew Energ 89: 680-694. doi: 10.1016/j.renene.2015.12.004
|
| [111] | Babonneau F, Haurie A, Vielle M (2014) Assessment of balanced burden-sharing in the 2050 EU climate/energy roadmap: a metamodeling approach. Climatic Change 134: 505-519. |
| [112] |
Jonsson DK, Johansson B, Månsson A, et al. (2015) Energy security matters in the EU Energy Roadmap. Energ Strategy Rev 6: 48-56. doi: 10.1016/j.esr.2015.03.002
|
| [113] |
Turnyanskiy M, Neu R, Albanese R, et al. (2015) European roadmap to the realization of fusion energy: Mission for solution on heat-exhaust systems. Fusion Eng Des 96-97: 361-64. doi: 10.1016/j.fusengdes.2015.04.041
|
| [114] |
Xiong W, Wang Y, Mathiesen BV, et al. (2015) Heat roadmap china: New heat strategy to reduce energy consumption towards 2030. Energy 81: 274-285. doi: 10.1016/j.energy.2014.12.039
|
| [115] |
Odenberger M, Kjärstad J, Johnsson F (2013) Prospects for CCS in the EU energy roadmap to 2050. Energ Procedia 37: 7573-7581. doi: 10.1016/j.egypro.2013.06.701
|
| [116] |
Jeffrey H, Sedgwick J, Robinson C (2013) Technology roadmaps: An evaluation of their success in the renewable energy sector. Technol Forecast Soc Change 80: 1015-1027. doi: 10.1016/j.techfore.2012.09.016
|
| [117] | Hey C (2012) Low-carbon and energy strategies for the EU. The European commission's roadmaps: A sound agenda for green economy? GAIA 21: 43-47. |
| [118] |
McDowall W (2012) Technology roadmaps for transition management: The case of hydrogen energy. Technol Forecast Soc Change 79: 530-542. doi: 10.1016/j.techfore.2011.10.002
|
| [119] | Wang YD, Chen WM, Park YK (2011) Integrated regional Energ Policy and planning framework: Its application to the evaluation of the renewable roadmap of carbon free Jeju Island in South Korea. Chapter 13 In: Sustainable Systems and Energy Management at the Regional Level: Comparative Approaches. IGI Global Eds.: 236-260. |
| [120] |
Gómez A, Zubizarreta J, Dopazo C, et al. (2011) Spanish energy roadmap to 2020: Socioeconomic implications of renewable targets. Energy 36: 1973-1985. doi: 10.1016/j.energy.2010.02.046
|
| [121] |
Amer M, Daim TU (2010) Application of technology roadmaps for renewable energy sector. Technol Forecast Soc Change 77: 1355-1370. doi: 10.1016/j.techfore.2010.05.002
|
| [122] |
Pang X, Mörtberg U, Brown N (2014) Energy models from a strategic environmental assessment perspective in an EU context - What is missing concerning renewables? Renew Sust Energ Rev 33: 353-362. doi: 10.1016/j.rser.2014.02.005
|
| [123] | Miran W, Nawaz M, Jang J, et al. (2016) Sustainable electricity generation by biodegradation of low-cost lemon peel biomass in a dual chamber microbial fuel cell. Int Biodeter Biodegr 106: 75-79. |
| [124] | He H, Zhou M, Yang J, et al. (2014).Simultaneous wastewater treatment, electricity generation and biomass production by an immobilized photosynthetic algal microbial fuel cell. Bioproc Biosyst Eng 37: 873-880. |
| [125] | Kondaveeti S, Choi KS, Kakarla R, et al. (2014) Microalgae Scenedesmus obliquus as renewable biomass feedstock for electricity generation in microbial fuel cells (MFCs). Front Environ Sci Eng 8: 784-791. |
| [126] |
Nishio K, Hashimoto K, Watanabe K (2013) Digestion of algal biomass for electricity generation in microbial fuel cells. Biosci Biotech Bioch 77: 670-672. doi: 10.1271/bbb.120833
|
| [127] |
Rashid N, Cui YF, Muhammad SUR, et al. (2013) Enhanced electricity generation by using algae biomass and activated sludge in microbial fuel cell. Sci Total Environ 456-457: 91-94. doi: 10.1016/j.scitotenv.2013.03.067
|
| [128] |
Zhang J, Zhang B, Tian C, et al. (2013) Simultaneous sulfide removal and electricity generation with corn stover biomass as co-substrate in microbial fuel cells. Bioresource Technol 138: 198-203. doi: 10.1016/j.biortech.2013.03.167
|
| [129] | Cheng H, Qian Q, Wang X, et al. (2012) Electricity generation from carboxymethyl cellulose biomass: A new application of enzymatic biofuel cells. Electrochim Acta 82: 203-207. |
| [130] |
Toonssen R, Woudstra N, Verkooijen AHM (2010) Decentralized generation of electricity with solid oxide fuel cells from centrally converted biomass. Int J Hydrogen Energ 35: 7594-7607. doi: 10.1016/j.ijhydene.2010.05.006
|
| [131] | Wang X, Feng Y, Wang H, et al. (2009) Bioaugmentation for electricity generation from corn stover biomass using microbial fuel cells. Environ Sci Technol 43: 6088-6093. |
| [132] |
Zhang Y, Min B, Huang L, et al. (2009) Generation of electricity and analysis of microbial communities in wheat straw biomass-powered microbial fuel cells. Appl Environ Microbiol 75: 3389-3395. doi: 10.1128/AEM.02240-08
|
| [133] | Macedo WN, Monteiro LG, Corgozinho IM, et al. (2016) Biomass based microturbine system for electricity generation for isolated communities in Amazon region. Renew Energ 91: 323-333. |
| [134] | Cerón AMR, Weingärtner S, Kafarov V (2015) Generation of electricity by plant biomass in villages of the Colombian provinces: Chocó, Meta and Putumayo. Chem Eng T 43: 577-582. |
| [135] | Umar MS, Jennings P, Urmee T (2014) Sustainable electricity generation from oil palm biomass wastes in Malaysia: An industry survey. Energy 67: 496-505. |
| [136] |
Moore S, Durant V, Mabee WE (2013) Determining appropriate feed-in tariff rates to promote biomass-to-electricity generation in Eastern Ontario, Canada. Energ Policy 63: 607-613. doi: 10.1016/j.enpol.2013.08.076
|
| [137] |
Rodriguez LC, May B, Herr A, et al. (2011) Biomass assessment and small scale biomass fired electricity generation in the Green Triangle, Australia. Biomass Bioenerg 35: 2589-2599. doi: 10.1016/j.biombioe.2011.02.030
|
| [138] |
Wei L, To SDF, Pordesimo L, et al. (2011) Evaluation of micro-scale electricity generation cost using biomass-derived synthetic gas through modelling. Int J Energ Res 35: 989-1003. doi: 10.1002/er.1749
|
| [139] | Evans A, Strezov V, Evans TJ (2010) Sustainability considerations for electricity generation from biomass. Renew Sust Energ Rev 14: 1419-1427. |
| [140] | Nasiri F, Zaccour G (2009) An exploratory game-theoretic analysis of biomass electricity generation supply chain. Energ Policy 37: 4514-4522. |
| [141] | Akgul O, Mac Dowell N, Papageorgiou LG, et al. (2014) A mixed integer nonlinear programming (MINLP) supply chain optimisation framework for carbon negative electricity generation using biomass to energy with CCS (BECCS) in the UK. Int J Greenhouse Gas Control 28: 189-202. |
| [142] |
Favero A, Massetti E (2014) Trade of woody biomass for electricity generation under climate mitigation policy. Resource Energ Econ 36: 166-190. doi: 10.1016/j.reseneeco.2013.11.005
|
| [143] | O’Shaughnessy SM, Deasy MJ, Kinsella CE, et al. (2013) Small scale electricity generation from a portable biomass cookstove: Prototype design and preliminary results. Appl Energ 102: 374-385. |
| [144] | Dwivedi P, Bailis R, Carter DR, et al. (2012) A landscape-based approach for assessing spatiotemporal impacts of forest biomass-based electricity generation on the age structure of surrounding forest plantations in the Southern United States. GCB Bioenergy 4: 342-357. |
| [145] |
Perilhon C, Alkadee D, Descombes G, et al. (2012) Life cycle assessment applied to electricity generation from renewable biomass. Energy Procedia 18: 165-176. doi: 10.1016/j.egypro.2012.05.028
|
| [146] | Shafie SM, Mahlia TMI, Masjuki HH, et al. (2012) A review on electricity generation based on biomass residue in Malaysia. Renew Sust Energ Rev 16: 5879-5889. |
| [147] | Basu P, Butler J, Leon MA (2011). Biomass co-firing options on the emission reduction and electricity generation costs in coal-fired power plants. Renew Energy 36: 282-288. |
| [148] | Hansson J, Berndes G, Johnsson F, et al. (2009) Co-firing biomass with coal for electricity generation-An assessment of the potential in EU27. Energ Policy 37: 1444-1455. |
| [149] | Berggren M, Ljunggren, E, Johnsson F (2008) Biomass co-firing potentials for electricity generation in Poland-Matching supply and co-firing opportunities. Biomass Bioenerg 32: 865-879. |
| [150] |
Panklib T, Prakasvudhisarn C, Khummongkol D (2014) The potential of biomass energy from agricultural residues for small electricity generation in Thailand. Energ Source 36: 803-814. doi: 10.1080/15567036.2010.545809
|
| [151] |
Skorek-Osikowska A, Bartela T, Kotowicz J, et al. (2014) The influence of the size of the CHP (combined heat and power) system integrated with a biomass fueled gas generator and piston engine on the thermodynamic and economic effectiveness of electricity and heat generation. Energy 67: 328-340. doi: 10.1016/j.energy.2014.01.015
|
| [152] |
Ruiz JA, Juárez MC, Morales MP, et al. (2013) Biomass gasification for electricity generation: Review of current technology barriers. Renew Sust Energ Rev 18: 174-183. doi: 10.1016/j.rser.2012.10.021
|
| [153] | Dasappa S (2011) Potential of biomass energy for electricity generation in sub-Saharan Africa. Energ Sust Dev 15: 203-213. |
| [154] | Liu X, Farmer M, Capareda S (2011) The Economic Feasibility of Electricity Generation from Biomass on the South Plains of Texas. In: The Economics of Alternative Energy Sources and Globalization. Edited by Andrew Schmitz, Norbert L. Wilson, Charles B. Moss, David Zilberman. Bentham Eds., 158-169. |
| [155] |
Pihl Erik E, Heyne S, Thunman H, et al. (2010) Highly efficient electricity generation from biomass by integration and hybridization with combined cycle gas turbine (CCGT) plants for natural gas. Energy 35: 4042-4052. doi: 10.1016/j.energy.2010.06.008
|
| [156] | Bazmi AA, Zahedi G, Hashim H (2011) Progress and challenges in utilization of palm oil biomass as fuel for decentralized electricity generation. Renew Sust Energ Rev 15: 574-583. |
| [157] | Hiloidhari M, Baruah DC (2011) Rice straw residue biomass potential for decentralized electricity generation: A GIS based study in Lakhimpur district of Assam, India. Energ Sust Dev 15: 214-222. |
| [158] | Kumar M, Patel SK, Mishra S (2010) Studies on characteristics of some shrubaceous non-woody biomass species and their electricity generation potentials. Energ Source 32: 786-795. |
| [159] |
Toonssen R, Woudstra N, Verkooijen AHM (2009) Decentralized generation of electricity from biomass with proton exchange membrane fuel cell. J Power Source 194: 456-466. doi: 10.1016/j.jpowsour.2009.05.044
|
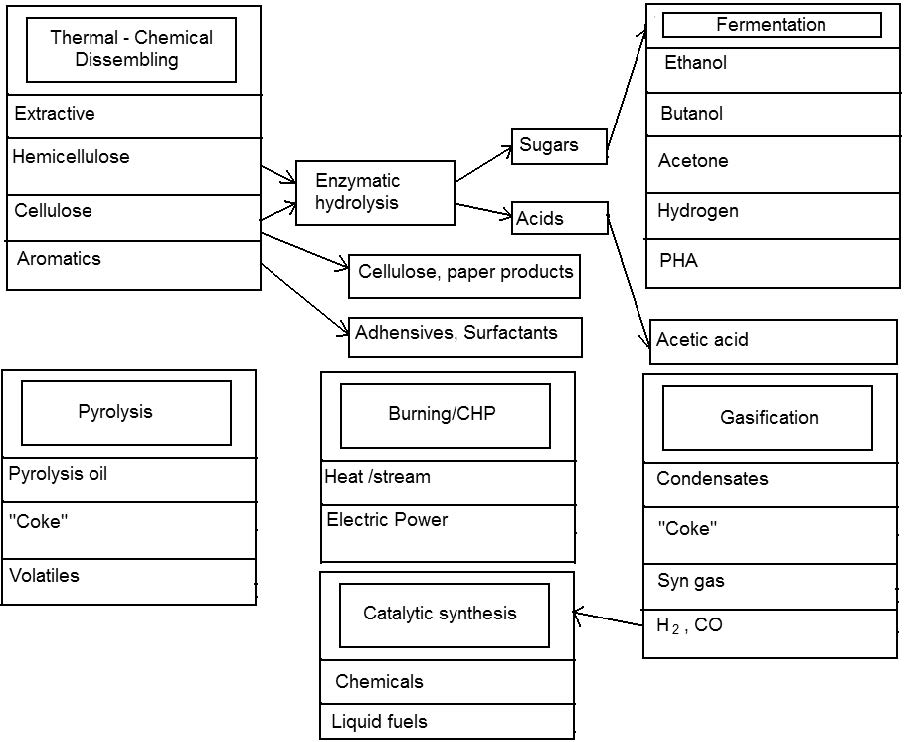
| [160] | Liu J, Deng Y (2008) The Biorefinery: Employing Biomass to Relieve Our Dependence on Fossil Sources. Biorefinery J Biobased Mater Bioenerg 2: 97-99. Available from: http://www.biorefineryresearchinstitute.com/biorefinery.php. |
| [161] | Anonymous (2010) Biomass – Assessing the Potential in Rural Areas. From: “Assessing Biomass Feasibility” project, which was undertaken as part of the University of Strathclyde’s Sustainable Engineering, MSc in “Energy Systems and the Environment”. Available from: http://www.esru.strath.ac.uk/EandE/Web_sites/09-10/Rural_renewables/Methodology_Specific Consideration_ Biomass_BiomassBoiler.html. |
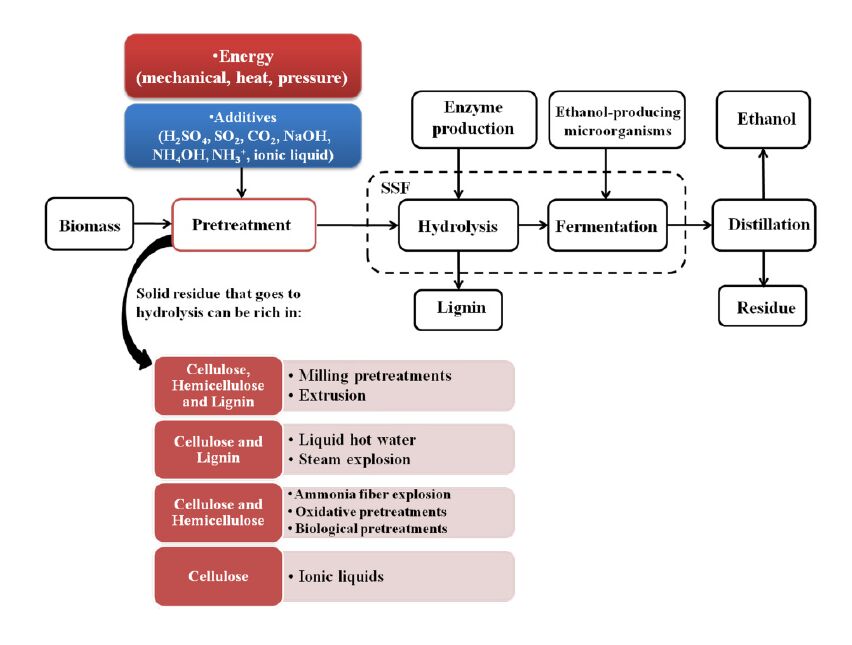
| [162] | Silva AS, Teixeira RSS, Moutta RO, et al. (2013) Sugarcane and Woody Biomass Pretreatments for Ethanol Production. Chapter 3 In: Sustainable Degradation of Lignocellulosic Biomass - Techniques, Applications and Commercialization. Edited by Anuj K. Chandel and Silvio Silvério da Silva. InTech Eds. 42pp. |
| [163] | Fatehi Pedram (2013) Production of Biofuels from Cellulose of Woody Biomass. Chapter 3 In: Cellulose - Biomass Conversion. Edited by Theo van de Ven and John Kadla, InTech Eds. Available from: http://www.intechopen.com/books/cellulose-biomass-conversion/production-of-biofuels-from-cellulose-of-woody-biomass. |
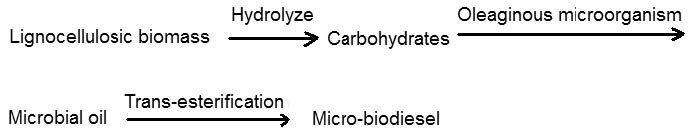
| [164] | Dai CC, Tao J, Wang Y, et al. (2010) Research advance and superiority of microdiesel production with biowastes. Afr J Microbiol Res 4: 977-983. |
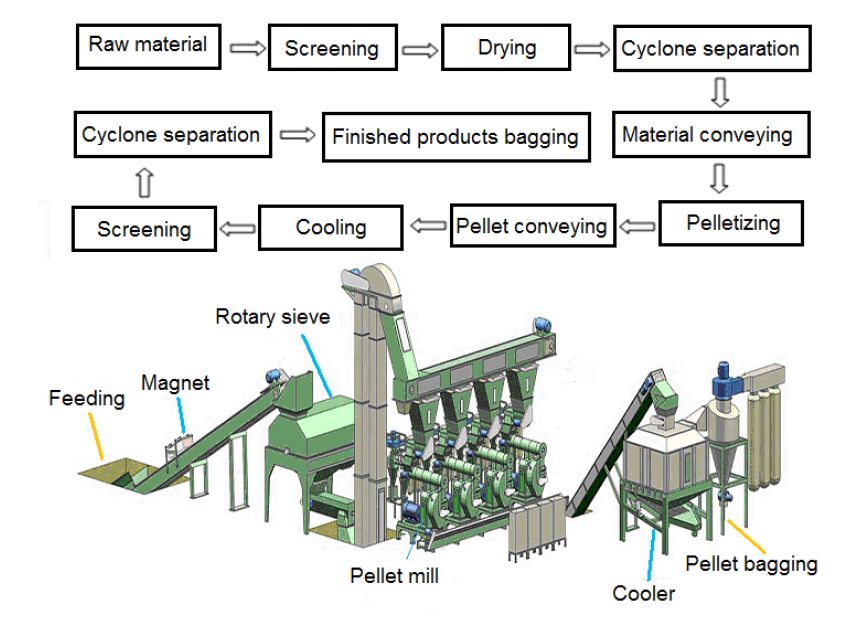
| [165] | Huang J (2015) Pellet Plant Process Flow Chart: Gemco Energy. Available from: http://www.biomass-energy.org/blog/pellet-plant-process-flow-chart.html. |
| [166] |
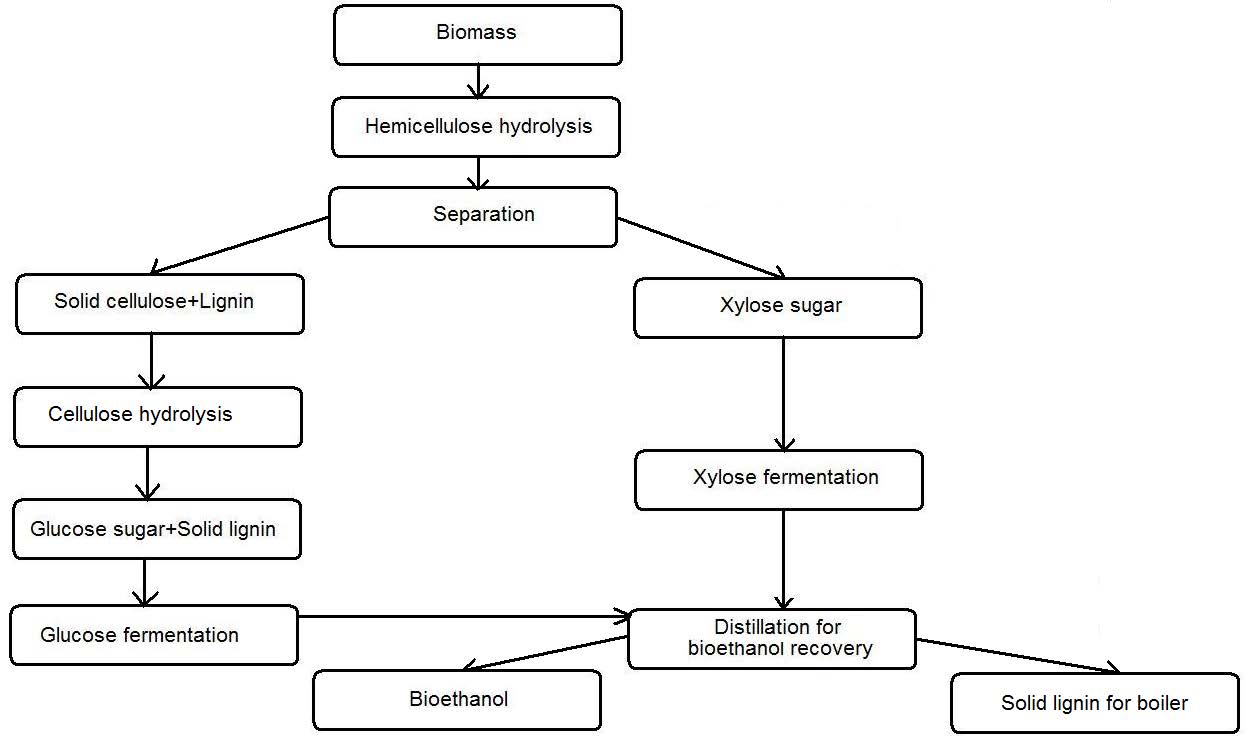
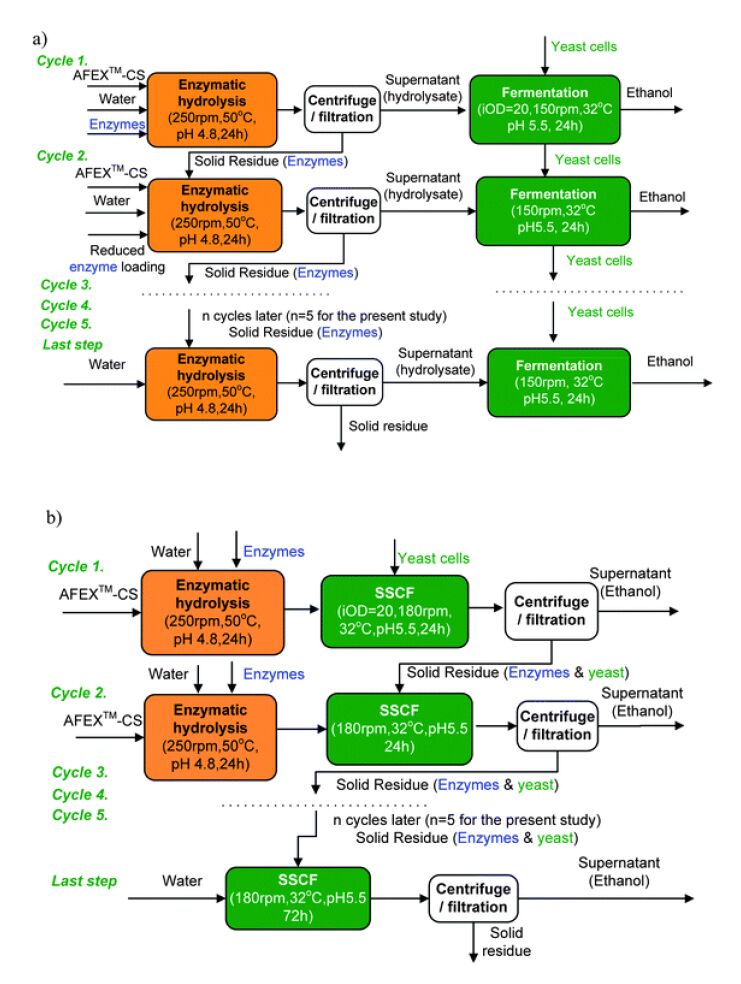
Jin M, Gunawan C, Uppugundla N, et al. (2012) A novel integrated biological process for cellulosic ethanol production featuring high ethanol productivity, enzyme recycling and yeast cells reuse. Energ Environ Sci 5: 7168-7175. doi: 10.1039/c2ee03058f
|

Figures(7) / Tables(10)

Grigorios L. Kyriakopoulos, Garyfallos Arabatzis, Miltiadis Chalikias. Renewables exploitation for energy production and biomass use for electricity generation. A multi-parametric literature-based review[J]. AIMS Energy, 2016, 4(5): 762-803. doi: 10.3934/energy.2016.5.762







 DownLoad:
DownLoad: