Citation: Murtaza Shabbir Hussain, Gabriel M Rodriguez, Difeng Gao, Michael Spagnuolo, Lauren Gambill, Mark Blenner. Recent advances in bioengineering of the oleaginous yeast Yarrowia lipolytica[J]. AIMS Bioengineering, 2016, 3(4): 493-514. doi: 10.3934/bioeng.2016.4.493

| [1] | Goncalves FA, Colen G, Takahashi JA (2014) Yarrowia lipolytica and its multiple applications in the biotechnological industry. Sci World J 2014: 476207. |
| [2] |
Zhu Q, Jackson EN (2015) Metabolic engineering of Yarrowia lipolytica for industrial applications. Curr Opin Biotechnol 36: 65–72. doi: 10.1016/j.copbio.2015.08.010

|
| [3] |
Cordero Otero R, Gaillardin C (1996) Efficient selection of hygromycin-B-resistant Yarrowia lipolytica transformants. Appl Microbiol Biotechnol 46: 143–148. doi: 10.1007/s002530050796
|
| [4] |
Chen DC, Beckerich JM, Gaillardin C (1997) One-step transformation of the dimorphic yeast Yarrowia lipolytica. Appl Microbiol Biotechnol 48: 232–235. doi: 10.1007/s002530051043
|
| [5] | Liu L, Otoupal P, Pan A, et al. (2014) Increasing expression level and copy number of a Yarrowia lipolytica plasmid through regulated centromere function. FEMS Yeast Res 14: 1124–1127. |
| [6] |
Hussain MS, Gambill L, Smith S, et al. (2016) Engineering promoter architecture in oleaginous yeast Yarrowia lipolytica. ACS Synth Biol 5: 213–223. doi: 10.1021/acssynbio.5b00100
|
| [7] |
Blazeck J, Reed B, Garg R, et al. (2013) Generalizing a hybrid synthetic promoter approach in Yarrowia lipolytica. Appl Microbiol Biotechnol 97: 3037–3052. doi: 10.1007/s00253-012-4421-5
|
| [8] |
Blazeck J, Liu L, Redden H, et al. (2011) Tuning gene expression in Yarrowia lipolytica by a hybrid promoter approach. Appl Environ Microbiol 77: 7905–7914. doi: 10.1128/AEM.05763-11
|
| [9] | Schwartz CM, Hussain MS, Blenner M, et al. (2016) Synthetic RNA polymerase III promoters facilitate high-efficiency CRISPR-Cas9-mediated genome editing in Yarrowia lipolytica. ACS Synth Biol. |
| [10] | Blazeck J, Hill A, Liu L, et al. (2014) Harnessing Yarrowia lipolytica lipogenesis to create a platform for lipid and biofuel production. Nat Commun 5: 3131. |
| [11] |
Tai M, Stephanopoulos G (2013) Engineering the push and pull of lipid biosynthesis in oleaginous yeast Yarrowia lipolytica for biofuel production. Metab Eng 15: 1–9. doi: 10.1016/j.ymben.2012.08.007
|
| [12] |
Qiao K, Imam Abidi SH, Liu H, et al. (2015) Engineering lipid overproduction in the oleaginous yeast Yarrowia lipolytica. Metab Eng 29: 56–65. doi: 10.1016/j.ymben.2015.02.005
|
| [13] |
Xu P, Qiao KJ, Ahn WS, et al. (2016) Engineering Yarrowia lipolytica as a platform for synthesis of drop-in transportation fuels and oleochemicals. P Natl Acad Sci USA 113: 10848–10853. doi: 10.1073/pnas.1607295113
|
| [14] |
Friedlander J, Tsakraklides V, Kamineni A, et al. (2016) Engineering of a high lipid producing Yarrowia lipolytica strain. Biotechnol Biofuels 9: 77. doi: 10.1186/s13068-016-0492-3
|
| [15] |
Xue Z, Sharpe PL, Hong SP, et al. (2013) Production of omega-3 eicosapentaenoic acid by metabolic engineering of Yarrowia lipolytica. Nat Biotechnol 31: 734–740. doi: 10.1038/nbt.2622
|
| [16] |
Smit MS, Mokgoro MM, Setati E, et al. (2005) alpha,omega-Dicarboxylic acid accumulation by acyl-CoA oxidase deficient mutants of Yarrowia lipolytica. Biotechnol Lett 27: 859–864. doi: 10.1007/s10529-005-6719-1
|
| [17] |
Haddouche R, Poirier Y, Delessert S, et al. (2011) Engineering polyhydroxyalkanoate content and monomer composition in the oleaginous yeast Yarrowia lipolytica by modifying the ss-oxidation multifunctional protein. Appl Microbiol Biotechnol 91: 1327–1340. doi: 10.1007/s00253-011-3331-2
|
| [18] |
Blazeck J, Hill A, Jamoussi M, et al. (2015) Metabolic engineering of Yarrowia lipolytica for itaconic acid production. Metab Eng 32: 66–73. doi: 10.1016/j.ymben.2015.09.005
|
| [19] |
Ledesma-Amaro R, Dulermo R, Niehus X, et al. (2016) Combining metabolic engineering and process optimization to improve production and secretion of fatty acids. Metab Eng 38: 38–46. doi: 10.1016/j.ymben.2016.06.004
|
| [20] |
Zhou J, Yin X, Madzak C, et al. (2012) Enhanced alpha-ketoglutarate production in Yarrowia lipolytica WSH-Z06 by alteration of the acetyl-CoA metabolism. J Biotechnol 161: 257–264. doi: 10.1016/j.jbiotec.2012.05.025
|
| [21] |
Yovkova V, Otto C, Aurich A, et al. (2014) Engineering the alpha-ketoglutarate overproduction from raw glycerol by overexpression of the genes encoding NADP+-dependent isocitrate dehydrogenase and pyruvate carboxylase in Yarrowia lipolytica. Appl Microbiol Biotechnol 98: 2003–2013. doi: 10.1007/s00253-013-5369-9
|
| [22] |
Wang G, Xiong X, Ghogare R, et al. (2016) Exploring fatty alcohol-producing capability of Yarrowia lipolytica. Biotechnol Biofuels 9: 107. doi: 10.1186/s13068-016-0512-3
|
| [23] |
Rutter CD, Rao CV (2016) Production of 1-decanol by metabolically engineered Yarrowia lipolytica. Metab Eng 38: 139–147. doi: 10.1016/j.ymben.2016.07.011
|
| [24] |
Wang W, Wei H, Knoshaug E, et al. (2016) Fatty alcohol production in Lipomyces starkeyi and Yarrowia lipolytica. Biotechnol Biofuels 9: 227. doi: 10.1186/s13068-016-0647-2
|
| [25] | Ryu S, Hipp J, Trinh CT (2015) Activating and elucidating metabolism of complex sugars in Yarrowia lipolytica. Appl Environ Microbiol 82: 1334–1345. |
| [26] |
Rodriguez GM, Hussain MS, Gambill L, et al. (2016) Engineering xylose utilization in Yarrowia lipolytica by understanding its cryptic xylose pathway. Biotechnol Biofuels 9: 149. doi: 10.1186/s13068-016-0562-6
|
| [27] |
Ledesma-Amaro R, Nicaud JM (2016) Metabolic engineering for expanding the substrate range of Yarrowia lipolytica. Trends Biotechnol 34: 798–809. doi: 10.1016/j.tibtech.2016.04.010
|
| [28] |
Li H, Alper HS (2016) Enabling xylose utilization in Yarrowia lipolytica for lipid production. Biotechnol J 11: 1230–1240. doi: 10.1002/biot.201600210
|
| [29] |
Ledesma-Amaro R, Lazar Z, Rakicka M, et al. (2016) Metabolic engineering of Yarrowia lipolytica to produce chemicals and fuels from xylose. Metab Eng 38: 115–124. doi: 10.1016/j.ymben.2016.07.001
|
| [30] |
Ledesma-Amaro R, Dulermo T, Nicaud JM (2015) Engineering Yarrowia lipolytica to produce biodiesel from raw starch. Biotechnol Biofuels 8: 148. doi: 10.1186/s13068-015-0335-7
|
| [31] |
Celinska E, Bialas W, Borkowska M, et al. (2015) Cloning, expression, and purification of insect (Sitophilus oryzae) alpha-amylase, able to digest granular starch, in Yarrowia lipolytica host. Appl Microbiol Biotechnol 99: 2727–2739. doi: 10.1007/s00253-014-6314-2
|
| [32] |
Yang CH, Huang YC, Chen CY, et al. (2010) Heterologous expression of Thermobifida fusca thermostable alpha-amylase in Yarrowia lipolytica and its application in boiling stable resistant sago starch preparation. J Ind Microbiol Biotechnol 37: 953–960. doi: 10.1007/s10295-010-0745-2
|
| [33] |
Xu J, Xiong P, He B (2016) Advances in improving the performance of cellulase in ionic liquids for lignocellulose biorefinery. Bioresour Technol 200: 961–970. doi: 10.1016/j.biortech.2015.10.031
|
| [34] |
Dujon B, Sherman D, Fischer G, et al. (2004) Genome evolution in yeasts. Nature 430: 35–44. doi: 10.1038/nature02579
|
| [35] | Liu L, Alper HS (2014) Draft genome sequence of the oleaginous yeast Yarrowia lipolytica PO1f, a commonly used metabolic engineering host. Genome Announc 2. |
| [36] |
Magnan C, Yu J, Chang I, et al. (2016) Sequence assembly of Yarrowia lipolytica strain W29/CLIB89 shows transposable element diversity. PLoS One 11: e0162363. doi: 10.1371/journal.pone.0162363
|
| [37] |
van Heerikhuizen H, Ykema A, Klootwijk J, et al. (1985) Heterogeneity in the ribosomal RNA genes of the yeast Yarrowia lipolytica; cloning and analysis of two size classes of repeats. Gene 39: 213–222. doi: 10.1016/0378-1119(85)90315-4
|
| [38] |
Davidow LS, Apostolakos D, O’Donnell MM, et al. (1985) Integrative transformation of the yeast Yarrowia lipolytica. Curr Genet 10: 39–48. doi: 10.1007/BF00418492
|
| [39] | Barth G, Gaillardin C (1996) Yarrowia lipolytica, In: Wolf K, Nonconventional Yeasts in Biotechnology, NewYork: Springer. |
| [40] |
Wang JH, Hung W, Tsai SH (2011) High efficiency transformation by electroporation of Yarrowia lipolytica. J Microbiol 49: 469–472. doi: 10.1007/s12275-011-0433-6
|
| [41] |
Duquesne S, Bordes F, Fudalej F, et al. (2012) The yeast Yarrowia lipolytica as a generic tool for molecular evolution of enzymes. Methods Mol Biol 861: 301–312. doi: 10.1007/978-1-61779-600-5_18
|
| [42] |
Theerachat M, Emond S, Cambon E, et al. (2012) Engineering and production of laccase from Trametes versicolor in the yeast Yarrowia lipolytica. Bioresour Technol 125: 267–274. doi: 10.1016/j.biortech.2012.07.117
|
| [43] |
Juretzek T, Wang HJ, Nicaud JM, et al. (2000) Comparison of promoters suitable for regulated overexpression of β-galactosidase in the alkane-utilizing yeast Yarrowia lipolytica. Biotechnol Bioproc Eng 5: 320–326. doi: 10.1007/BF02942206
|
| [44] | Wang HJ, Le Dall MT, Wach Y, et al. (1999) Evaluation of acyl coenzyme A oxidase (Aox) isozyme function in the n-alkane-assimilating yeast Yarrowia lipolytica. J Bacteriol 181: 5140–5148. |
| [45] |
Yamagami S, Morioka D, Fukuda R, et al. (2004) A basic helix-loop-helix transcription factor essential for cytochrome p450 induction in response to alkanes in yeast Yarrowia lipolytica. J Biol Chem 279: 22183–22189. doi: 10.1074/jbc.M313313200
|
| [46] |
Blanchin-Roland S, Cordero Otero RR, Gaillardin C (1994) Two upstream activation sequences control the expression of the XPR2 gene in the yeast Yarrowia lipolytica. Mol Cell Biol 14: 327–338. doi: 10.1128/MCB.14.1.327
|
| [47] | Madzak C, Blanchin-Roland S, Cordero Otero RR, et al. (1999) Functional analysis of upstream regulating regions from the Yarrowia lipolytica XPR2 promoter. Microbiology 145 (Pt 1): 75–87. |
| [48] | Madzak C, Treton B, Blanchin-Roland S (2000) Strong hybrid promoters and integrative expression/secretion vectors for quasi-constitutive expression of heterologous proteins in the yeast Yarrowia lipolytica. J Mol Microbiol Biotechnol 2: 207–216. |
| [49] |
Shabbir Hussain M, Gambill L, Smith S, et al. (2016) Engineering promoter architecture in oleaginous yeast Yarrowia lipolytica. ACS Synth Biol 5: 213–223. doi: 10.1021/acssynbio.5b00100
|
| [50] | Muller S, Sandal T, Kamp-Hansen P, et al. (1998) Comparison of expression systems in the yeasts Saccharomyces cerevisiae, Hansenula polymorpha, Klyveromyces lactis, Schizosaccharomyces pombe and Yarrowia lipolytica. Cloning of two novel promoters from Yarrowia lipolytica. Yeast 14: 1267–1283. |
| [51] |
Kozak M (1984) Compilation and analysis of sequences upstream from the translational start site in eukaryotic mRNAs. Nucleic Acids Res 12: 857–872. doi: 10.1093/nar/12.2.857
|
| [52] |
Kozak M (1986) Point mutations define a sequence flanking the AUG initiator codon that modulates translation by eukaryotic ribosomes. Cell 44: 283–292. doi: 10.1016/0092-8674(86)90762-2
|
| [53] | Merkulov S, van Assema F, Springer J, et al. (2000) Cloning and characterization of the Yarrowia lipolytica squalene synthase (SQS1) gene and functional complementation of the Saccharomyces cerevisiae erg9 mutation. Yeast 16: 197–206. |
| [54] |
Zhang B, Rong C, Chen H, et al. (2012) De novo synthesis of trans-10, cis-12 conjugated linoleic acid in oleaginous yeast Yarrowia lipolytica. Microb Cell Fact 11: 1–8. doi: 10.1186/1475-2859-11-1
|
| [55] |
Kramara J, Willcox S, Gunisova S, et al. (2010) Tay1 protein, a novel telomere binding factor from Yarrowia lipolytica. J Biol Chem 285: 38078–38092. doi: 10.1074/jbc.M110.127605
|
| [56] | Farrel C, Mayorga M, Chevreux B (2013) Carotene hydroxylase and its use for producing carotenoids. In: Ltd DNP. |
| [57] |
Gasmi N, Fudalej F, Kallel H, et al. (2011) A molecular approach to optimize hIFN alpha2b expression and secretion in Yarrowia lipolytica. Appl Microbiol Biotechnol 89: 109–119. doi: 10.1007/s00253-010-2803-0
|
| [58] | Nakagawa S, Niimura Y, Gojobori T, et al. (2008) Diversity of preferred nucleotide sequences around the translation initiation codon in eukaryote genomes. Nucleic Acids Res 36: 861–871. |
| [59] |
Schwartz CM, Hussain MS, Blenner M, et al. (2016) Synthetic RNA Polymerase III Promoters Facilitate High-Efficiency CRISPR-Cas9-Mediated Genome Editing in Yarrowia lipolytica. ACS Synth Biol 5: 356–359. doi: 10.1021/acssynbio.5b00162
|
| [60] |
Gao S, Han L, Zhu L, et al. (2014) One-step integration of multiple genes into the oleaginous yeast Yarrowia lipolytica. Biotechnol Lett 36: 2523–2528. doi: 10.1007/s10529-014-1634-y
|
| [61] |
Curran KA, Morse NJ, Markham KA, et al. (2015) Short synthetic terminators for improved heterologous gene expression in yeast. ACS Synth Biol 4: 824–832. doi: 10.1021/sb5003357
|
| [62] |
Laine JP, Singh BN, Krishnamurthy S, et al. (2009) A physiological role for gene loops in yeast. Genes Dev 23: 2604–2609. doi: 10.1101/gad.1823609
|
| [63] |
Kanaar R, Hoeijmakers JH, van Gent DC (1998) Molecular mechanisms of DNA double strand break repair. Trends Cell Biol 8: 483–489. doi: 10.1016/S0962-8924(98)01383-X
|
| [64] |
Pastink A, Lohman PH (1999) Repair and consequences of double-strand breaks in DNA. Mutat Res 428: 141–156. doi: 10.1016/S1383-5742(99)00042-3
|
| [65] | Pastink A, Eeken JC, Lohman PH (2001) Genomic integrity and the repair of double-strand DNA breaks. Mutat Res 480: 37–50. |
| [66] |
Loeillet S, Palancade B, Cartron M, et al. (2005) Genetic network interactions among replication, repair and nuclear pore deficiencies in yeast. DNA Repair (Amst) 4: 459–468. doi: 10.1016/j.dnarep.2004.11.010
|
| [67] |
Fickers P, Le Dall MT, Gaillardin C, et al. (2003) New disruption cassettes for rapid gene disruption and marker rescue in the yeast Yarrowia lipolytica. J Microbiol Methods 55: 727–737. doi: 10.1016/j.mimet.2003.07.003
|
| [68] |
Van Dyck E, Stasiak AZ, Stasiak A, et al. (1999) Binding of double-strand breaks in DNA by human Rad52 protein. Nature 398: 728–731. doi: 10.1038/19560
|
| [69] |
Verbeke J, Beopoulos A, Nicaud JM (2013) Efficient homologous recombination with short length flanking fragments in Ku70 deficient Yarrowia lipolytica strains. Biotechnol Lett 35: 571–576. doi: 10.1007/s10529-012-1107-0
|
| [70] |
Kretzschmar A, Otto C, Holz M, et al. (2013) Increased homologous integration frequency in Yarrowia lipolytica strains defective in non-homologous end-joining. Curr Genet 59: 63–72. doi: 10.1007/s00294-013-0389-7
|
| [71] | Vasiliki T, Elena B, Gregory S, et al. (2015) Improved gene targeting through cell cycle synchronization. PLoS One 10. |
| [72] | Isaac RS, Jiang F, Doudna JA, et al. (2016) Nucleosome breathing and remodeling constrain CRISPR-Cas9 function. Elife 5. |
| [73] | Daer R, Cutts JP, Brafman DA, et al. (2016) The impact of chromatin dynamics on Cas9-mediated genome editing in human cells. ACS Synth Biol. |
| [74] | Hamilton DL, Abremski K (1984) Site-specific recombination by the bacteriophage P1 lox-Cre system. Cre-mediated synapsis of two lox sites. J Mol Biol 178: 481–486. |
| [75] |
Le Dall MT, Nicaud JM, Gaillardin C (1994) Multiple-copy integration in the yeast Yarrowia lipolytica. Curr Genet 26: 38–44. doi: 10.1007/BF00326302
|
| [76] |
Casaregola S, Feynerol C, Diez M, et al. (1997) Genomic organization of the yeast Yarrowia lipolytica. Chromosoma 106: 380–390. doi: 10.1007/s004120050259
|
| [77] |
Schmid-Berger N, Schmid B, Barth G (1994) Ylt1, a highly repetitive retrotransposon in the genome of the dimorphic fungus Yarrowia lipolytica. J Bacteriol 176: 2477–2482. doi: 10.1128/jb.176.9.2477-2482.1994
|
| [78] |
Mauersberger S, Wang HJ, Gaillardin C, et al. (2001) Insertional mutagenesis in the n-alkane-assimilating yeast Yarrowia lipolytica: generation of tagged mutations in genes involved in hydrophobic substrate utilization. J Bacteriol 183: 5102–5109. doi: 10.1128/JB.183.17.5102-5109.2001
|
| [79] |
Bordes F, Fudalej F, Dossat V, et al. (2007) A new recombinant protein expression system for high-throughput screening in the yeast Yarrowia lipolytica. J Microbiol Meth 70: 493–502. doi: 10.1016/j.mimet.2007.06.008
|
| [80] |
Pignede G, Wang HJ, Fudalej F, et al. (2000) Autocloning and amplification of LIP2 in Yarrowia lipolytica. Appl Environ Microbiol 66: 3283–3289. doi: 10.1128/AEM.66.8.3283-3289.2000
|
| [81] | Nicaud JM, Madzak C, van den Broek P, et al. (2002) Protein expression and secretion in the yeast Yarrowia lipolytica. FEMS Yeast Res 2: 371–379. |
| [82] |
Gao C, Qi Q, Madzak C, et al. (2015) Exploring medium-chain-length polyhydroxyalkanoates production in the engineered yeast Yarrowia lipolytica. J Ind Microbiol Biotechnol 42: 1255–1262. doi: 10.1007/s10295-015-1649-y
|
| [83] |
Ratledge C (2002) Regulation of lipid accumulation in oleaginous micro-organisms. Biochem Soc Trans 30: 1047–1050. doi: 10.1042/bst0301047
|
| [84] |
Beopoulos A, Cescut J, Haddouche R, et al. (2009) Yarrowia lipolytica as a model for bio-oil production. Prog Lipid Res 48: 375–387. doi: 10.1016/j.plipres.2009.08.005
|
| [85] |
Ratledge C (2014) The role of malic enzyme as the provider of NADPH in oleaginous microorganisms: a reappraisal and unsolved problems. Biotechnol Lett 36: 1557–1568. doi: 10.1007/s10529-014-1532-3
|
| [86] |
Wynn JP, Kendrick A, Hamid AA, et al. (1997) Malic enzyme: a lipogenic enzyme in fungi. Biochem Soc Trans 25: S669. doi: 10.1042/bst025s669
|
| [87] |
Zhang H, Zhang L, Chen H, et al. (2013) Regulatory properties of malic enzyme in the oleaginous yeast, Yarrowia lipolytica, and its non-involvement in lipid accumulation. Biotechnol Lett 35: 2091–2098. doi: 10.1007/s10529-013-1302-7
|
| [88] |
Wasylenko TM, Ahn WS, Stephanopoulos G (2015) The oxidative pentose phosphate pathway is the primary source of NADPH for lipid overproduction from glucose in Yarrowia lipolytica. Metab Eng 30: 27–39. doi: 10.1016/j.ymben.2015.02.007
|
| [89] |
Liu N, Qiao K, Stephanopoulos G (2016) 13C Metabolic Flux Analysis of acetate conversion to lipids by Yarrowia lipolytica. Metab Eng 38: 86–97. doi: 10.1016/j.ymben.2016.06.006
|
| [90] |
Morin N, Cescut J, Beopoulos A, et al. (2011) Transcriptomic analyses during the transition from biomass production to lipid accumulation in the oleaginous yeast Yarrowia lipolytica. PLoS One 6: e27966. doi: 10.1371/journal.pone.0027966
|
| [91] |
Beopoulos A, Mrozova Z, Thevenieau F, et al. (2008) Control of lipid accumulation in the yeast Yarrowia lipolytica. Appl Environ Microbiol 74: 7779–7789. doi: 10.1128/AEM.01412-08
|
| [92] | Beopoulos A, Haddouche R, Kabran P, et al. (2012) Identification and characterization of DGA2, an acyltransferase of the DGAT1 acyl-CoA:diacylglycerol acyltransferase family in the oleaginous yeast Yarrowia lipolytica. New insights into the storage lipid metabolism of oleaginous yeasts. Appl Microbiol Biotechnol 93: 1523–1537. |
| [93] |
Wang ZP, Xu HM, Wang GY, et al. (2013) Disruption of the MIG1 gene enhances lipid biosynthesis in the oleaginous yeast Yarrowia lipolytica ACA-DC 50109. Biochim Biophys Acta 1831: 675–682. doi: 10.1016/j.bbalip.2012.12.010
|
| [94] |
Seip J, Jackson R, He H, et al. (2013) Snf1 is a regulator of lipid accumulation in Yarrowia lipolytica. Appl Environ Microbiol 79: 7360–7370. doi: 10.1128/AEM.02079-13
|
| [95] |
Dulermo T, Nicaud JM (2011) Involvement of the G3P shuttle and beta-oxidation pathway in the control of TAG synthesis and lipid accumulation in Yarrowia lipolytica. Metab Eng 13: 482–491. doi: 10.1016/j.ymben.2011.05.002
|
| [96] |
Liu L, Pan A, Spofford C, et al. (2015) An evolutionary metabolic engineering approach for enhancing lipogenesis in Yarrowia lipolytica. Metab Eng 29: 36–45. doi: 10.1016/j.ymben.2015.02.003
|
| [97] |
Liu L, Markham K, Blazeck J, et al. (2015) Surveying the lipogenesis landscape in Yarrowia lipolytica through understanding the function of a Mga2p regulatory protein mutant. Metab Eng 31: 102–111. doi: 10.1016/j.ymben.2015.07.004
|
| [98] |
Poirier Y, Antonenkov VD, Glumoff T, et al. (2006) Peroxisomal beta-oxidation—a metabolic pathway with multiple functions. Biochim Biophys Acta 1763: 1413–1426. doi: 10.1016/j.bbamcr.2006.08.034
|
| [99] |
Dulermo R, Gamboa-Melendez H, Ledesma-Amaro R, et al. (2015) Unraveling fatty acid transport and activation mechanisms in Yarrowia lipolytica. Biochim Biophys Acta 1851: 1202–1217. doi: 10.1016/j.bbalip.2015.04.004
|
| [100] |
Haddouche R, Delessert S, Sabirova J, et al. (2010) Roles of multiple acyl-CoA oxidases in the routing of carbon flow towards beta-oxidation and polyhydroxyalkanoate biosynthesis in Yarrowia lipolytica. FEMS Yeast Res 10: 917–927. doi: 10.1111/j.1567-1364.2010.00670.x
|
| [101] |
Hazer B, Steinbuchel A (2007) Increased diversification of polyhydroxyalkanoates by modification reactions for industrial and medical applications. Appl Microbiol Biotechnol 74: 1–12. doi: 10.1007/s00253-006-0732-8
|
| [102] |
Rehm BH (2003) Polyester synthases: natural catalysts for plastics. Biochem J 376: 15–33. doi: 10.1042/bj20031254
|
| [103] |
Sabirova JS, Haddouche R, Van Bogaert I, et al. (2011) The “LipoYeasts” project: using the oleaginous yeast Yarrowia lipolytica in combination with specific bacterial genes for the bioconversion of lipids, fats and oils into high-value products. Microb Biotechnol 4: 47–54. doi: 10.1111/j.1751-7915.2010.00187.x
|
| [104] |
Chuah JA, Tomizawa S, Yamada M, et al. (2013) Characterization of site-specific mutations in a short-chain-length/medium-chain-length polyhydroxyalkanoate synthase: in vivo and in vitro studies of enzymatic activity and substrate specificity. Appl Environ Microbiol 79: 3813–3821. doi: 10.1128/AEM.00564-13
|
| [105] |
Tsigie YA, Wang CY, Truong CT, et al. (2011) Lipid production from Yarrowia lipolytica Po1g grown in sugarcane bagasse hydrolysate. Bioresour Technol 102: 9216–9222. doi: 10.1016/j.biortech.2011.06.047
|
| [106] |
Ruiz-Herrera J, Sentandreu R (2002) Different effectors of dimorphism in Yarrowia lipolytica. Arch Microbiol 178: 477–483. doi: 10.1007/s00203-002-0478-3
|
| [107] | Fontes GC, Amaral PF, Nele M, et al. (2010) Factorial design to optimize biosurfactant production by Yarrowia lipolytica. J Biomed Biotechnol 2010: 821306. |
| [108] | Pan LX, Yang DF, Shao L, et al. (2009) Isolation of the oleaginous yeasts from the soil and studies of their lipid-producing capacities. Food Technol Biotechnol 47: 215–220. |
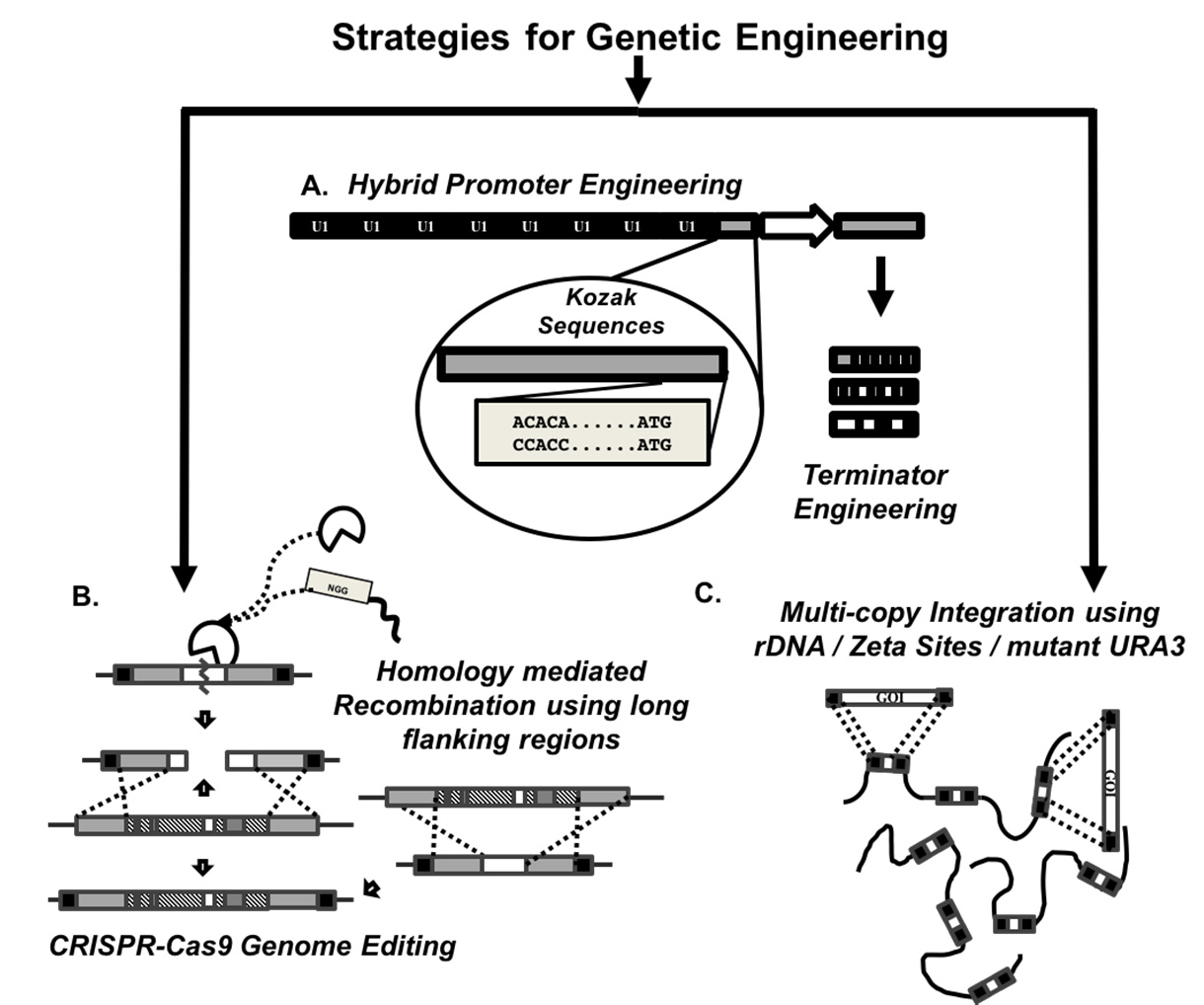
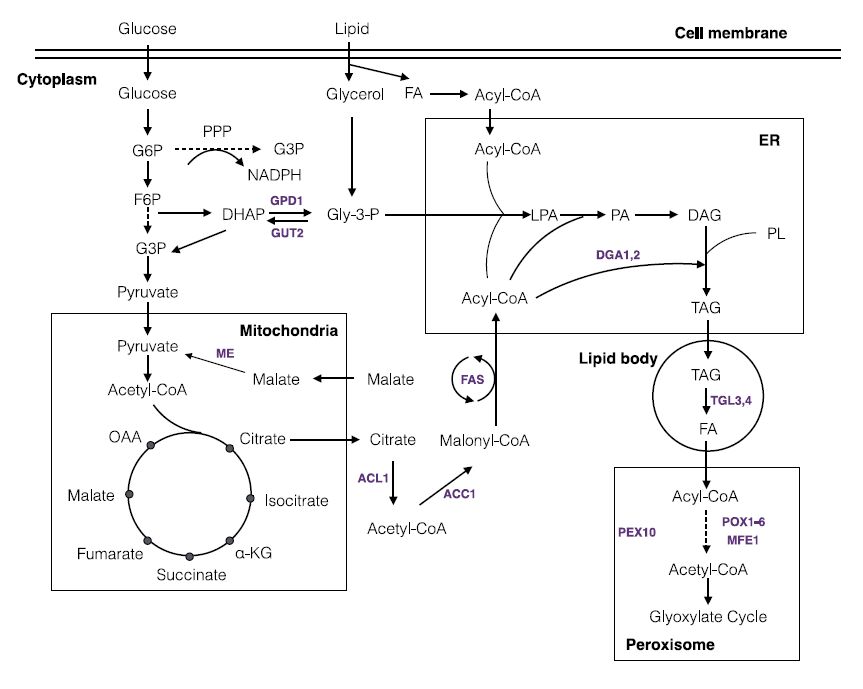
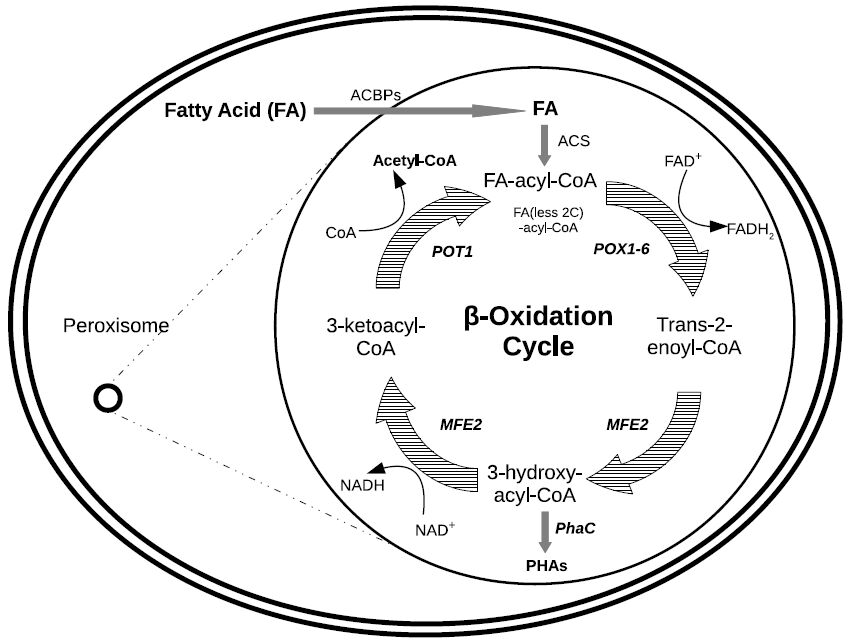
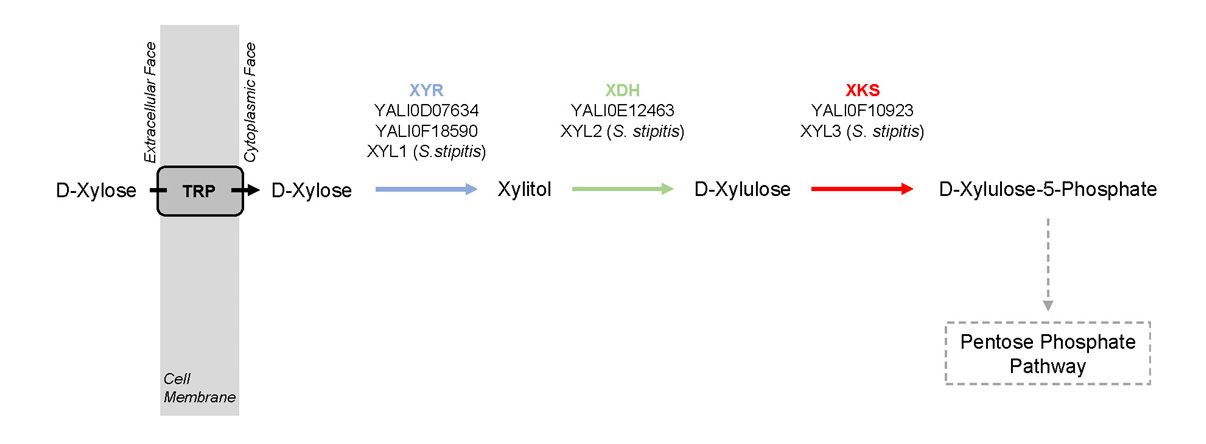
Figures(4) / Tables(2)

Murtaza Shabbir Hussain, Gabriel M Rodriguez, Difeng Gao, Michael Spagnuolo, Lauren Gambill, Mark Blenner. Recent advances in bioengineering of the oleaginous yeast Yarrowia lipolytica[J]. AIMS Bioengineering, 2016, 3(4): 493-514. doi: 10.3934/bioeng.2016.4.493







 DownLoad:
DownLoad: