Citation: Pawel Jajesniak, Tuck Seng Wong. From genetic circuits to industrial-scale biomanufacturing: bacterial promoters as a cornerstone of biotechnology[J]. AIMS Bioengineering, 2015, 2(3): 277-296. doi: 10.3934/bioeng.2015.3.277

| [1] |
Jacob F, Monod J (1961) Genetic regulatory mechanisms in the synthesis of proteins. J Mol Biol 3: 318-356. doi: 10.1016/S0022-2836(61)80072-7

|
| [2] |
Gorke B, Stulke J (2008) Carbon catabolite repression in bacteria: many ways to make the most out of nutrients. Nat Rev Microbiol 6: 613-624. doi: 10.1038/nrmicro1932
|
| [3] |
Terpe K (2006) Overview of bacterial expression systems for heterologous protein production: from molecular and biochemical fundamentals to commercial systems. Appl Microbiol Biotechnol 72: 211-222. doi: 10.1007/s00253-006-0465-8
|
| [4] |
Weickert MJ, Doherty DH, Best EA, et al. (1996) Optimization of heterologous protein production in Escherichia coli. Curr Opin Biotechnol 7: 494-499. doi: 10.1016/S0958-1669(96)80051-6
|
| [5] |
Blazeck J, Alper HS (2013) Promoter engineering: recent advances in controlling transcription at the most fundamental level. Biotechnol J 8: 46-58. doi: 10.1002/biot.201200120
|
| [6] | Busby S, Ebright RH (1994) Promoter structure, promoter recognition, and transcription activation in prokaryotes. Cell 79: 743-746. |
| [7] |
Hawley DK, McClure WR (1983) Compilation and analysis of Escherichia coli promoter DNA sequences. Nucleic Acids Res 11: 2237-2255. doi: 10.1093/nar/11.8.2237
|
| [8] | Harley CB, Reynolds RP (1987) Analysis of E. coli promoter sequences. Nucleic Acids Res 15: 2343-2361. |
| [9] | Oliphant AR, Struhl K (1988) Defining the consensus sequences of E.coli promoter elements by random selection. Nucleic Acids Res 16: 7673-7683. |
| [10] | Ishihama A (1993) Protein-protein communication within the transcription apparatus. J Bacteriol 175: 2483-2489. |
| [11] | Ebright RH, Cossart P, Gicquel-Sanzey B, et al. (1984) Mutations that alter the DNA sequence specificity of the catabolite gene activator protein of E. coli. Nature 311: 232-235. |
| [12] |
Lewis M, Chang G, Horton NC, et al. (1996) Crystal structure of the lactose operon repressor and its complexes with DNA and inducer. Science 271: 1247-1254. doi: 10.1126/science.271.5253.1247
|
| [13] |
Monsalve M, Calles B, Mencia M, et al. (1998) Binding of phage phi29 protein p4 to the early A2c promoter: recruitment of a repressor by the RNA polymerase. J Mol Biol 283: 559-569. doi: 10.1006/jmbi.1998.2084
|
| [14] |
Browning DF, Busby SJ (2004) The regulation of bacterial transcription initiation. Nat Rev Microbiol 2: 57-65. doi: 10.1038/nrmicro787
|
| [15] |
Balleza E, Lopez-Bojorquez LN, Martinez-Antonio A, et al. (2009) Regulation by transcription factors in bacteria: beyond description. FEMS Microbiol Rev 33: 133-151. doi: 10.1111/j.1574-6976.2008.00145.x
|
| [16] | Aoyama T, Takanami M, Ohtsuka E, et al. (1983) Essential structure of E. coli promoter: effect of spacer length between the two consensus sequences on promoter function. Nucleic Acids Res 11: 5855-5864. |
| [17] | Mulligan ME, Brosius J, McClure WR (1985) Characterization in vitro of the effect of spacer length on the activity of Escherichia coli RNA polymerase at the TAC promoter. J Biol Chem 260: 3529-3538. |
| [18] | Jensen PR, Hammer K (1998) The sequence of spacers between the consensus sequences modulates the strength of prokaryotic promoters. Appl Environ Microbiol 64: 82-87. |
| [19] |
Rud I, Jensen PR, Naterstad K, et al. (2006) A synthetic promoter library for constitutive gene expression in Lactobacillus plantarum. Microbiology 152: 1011-1019. doi: 10.1099/mic.0.28599-0
|
| [20] |
Alper H, Fischer C, Nevoigt E, et al. (2005) Tuning genetic control through promoter engineering. Proc Natl Acad Sci U S A 102: 12678-12683. doi: 10.1073/pnas.0504604102
|
| [21] |
de Boer HA, Comstock LJ, Vasser M (1983) The tac promoter: a functional hybrid derived from the trp and lac promoters. Proc Natl Acad Sci U S A 80: 21-25. doi: 10.1073/pnas.80.1.21
|
| [22] |
Yansura DG, Henner DJ (1984) Use of the Escherichia coli lac repressor and operator to control gene expression in Bacillus subtilis. Proc Natl Acad Sci U S A 81: 439-443. doi: 10.1073/pnas.81.2.439
|
| [23] | Haldimann A, Daniels LL, Wanner BL (1998) Use of new methods for construction of tightly regulated arabinose and rhamnose promoter fusions in studies of the Escherichia coli phosphate regulon. J Bacteriol 180: 1277-1286. |
| [24] |
Tawfik DS, Griffiths AD (1998) Man-made cell-like compartments for molecular evolution. Nat Biotechnol 16: 652-656. doi: 10.1038/nbt0798-652
|
| [25] |
Yonezawa M, Doi N, Kawahashi Y, et al. (2003) DNA display for in vitro selection of diverse peptide libraries. Nucleic Acids Res 31: e118. doi: 10.1093/nar/gng119
|
| [26] |
Levy M, Griswold KE, Ellington AD (2005) Direct selection of trans-acting ligase ribozymes by in vitro compartmentalization. RNA 11: 1555-1562. doi: 10.1261/rna.2121705
|
| [27] |
Ghadessy FJ, Ong JL, Holliger P (2001) Directed evolution of polymerase function by compartmentalized self-replication. Proc Natl Acad Sci U S A 98: 4552-4557. doi: 10.1073/pnas.071052198
|
| [28] |
Griffiths AD, Tawfik DS (2003) Directed evolution of an extremely fast phosphotriesterase by in vitro compartmentalization. EMBO J 22: 24-35. doi: 10.1093/emboj/cdg014
|
| [29] |
Griffiths AD, Tawfik DS (2006) Miniaturising the laboratory in emulsion droplets. Trends Biotechnol 24: 395-402. doi: 10.1016/j.tibtech.2006.06.009
|
| [30] |
Sepp A, Choo Y (2005) Cell-free selection of zinc finger DNA-binding proteins using in vitro compartmentalization. J Mol Biol 354: 212-219. doi: 10.1016/j.jmb.2005.09.051
|
| [31] |
Paul S, Stang A, Lennartz K, et al. (2013) Selection of a T7 promoter mutant with enhanced in vitro activity by a novel multi-copy bead display approach for in vitro evolution. Nucleic Acids Res 41: e29. doi: 10.1093/nar/gks940
|
| [32] |
Porro D, Gasser B, Fossati T, et al. (2011) Production of recombinant proteins and metabolites in yeasts: when are these systems better than bacterial production systems? Appl Microbiol Biotechnol 89: 939-948. doi: 10.1007/s00253-010-3019-z
|
| [33] |
Berlec A, Strukelj B (2013) Current state and recent advances in biopharmaceutical production in Escherichia coli, yeasts and mammalian cells. J Ind Microbiol Biotechnol 40: 257-274. doi: 10.1007/s10295-013-1235-0
|
| [34] |
Walsh G (2014) Biopharmaceutical benchmarks 2014. Nat Biotechnol 32: 992-1000. doi: 10.1038/nbt.3040
|
| [35] |
Westers L, Westers H, Quax WJ (2004) Bacillus subtilis as cell factory for pharmaceutical proteins: a biotechnological approach to optimize the host organism. Biochim Biophys Acta 1694: 299-310. doi: 10.1016/j.bbamcr.2004.02.011
|
| [36] |
Liu L, Yang H, Shin HD, et al. (2013) How to achieve high-level expression of microbial enzymes: strategies and perspectives. Bioengineered 4: 212-223. doi: 10.4161/bioe.24761
|
| [37] |
Retallack DM, Jin H, Chew L (2012) Reliable protein production in a Pseudomonas fluorescens expression system. Protein Expr Purif 81: 157-165. doi: 10.1016/j.pep.2011.09.010
|
| [38] | Chen R (2012) Bacterial expression systems for recombinant protein production: E. coli and beyond. Biotechnol Adv 30: 1102-1107. |
| [39] |
Sorensen HP, Mortensen KK (2005) Advanced genetic strategies for recombinant protein expression in Escherichia coli. J Biotechnol 115: 113-128. doi: 10.1016/j.jbiotec.2004.08.004
|
| [40] |
Gopal GJ, Kumar A (2013) Strategies for the production of recombinant protein in Escherichia coli. Protein J 32: 419-425. doi: 10.1007/s10930-013-9502-5
|
| [41] | Rosano GL, Ceccarelli EA (2014) Recombinant protein expression in Escherichia coli: advances and challenges. Front Microbiol 5: 172. |
| [42] | Pan SH, Malcolm BA (2000) Reduced background expression and improved plasmid stability with pET vectors in BL21 (DE3). Biotechniques 29: 1234-1238. |
| [43] |
Tegel H, Ottosson J, Hober S (2011) Enhancing the protein production levels in Escherichia coli with a strong promoter. FEBS J 278: 729-739. doi: 10.1111/j.1742-4658.2010.07991.x
|
| [44] |
Balzer S, Kucharova V, Megerle J, et al. (2013) A comparative analysis of the properties of regulated promoter systems commonly used for recombinant gene expression in Escherichia coli. Microb Cell Fact 12: 26. doi: 10.1186/1475-2859-12-26
|
| [45] |
Gronenborn B (1976) Overproduction of phage lambda repressor under control of the lac promotor of Escherichia coli. Mol Gen Genet 148: 243-250. doi: 10.1007/BF00332898
|
| [46] |
Silverstone AE, Arditti RR, Magasanik B (1970) Catabolite-insensitive revertants of lac promoter mutants. Proc Natl Acad Sci U S A 66: 773-779. doi: 10.1073/pnas.66.3.773
|
| [47] | Wanner BL, Kodaira R, Neidhardt FC (1977) Physiological regulation of a decontrolled lac operon. J Bacteriol 130: 212-222. |
| [48] | Bass SH, Yansura DG (2000) Application of the E. coli trp promoter. Mol Biotechnol 16: 253-260. |
| [49] |
Somerville RL (1988) The trp promoter of Escherichia coli and its exploitation in the design of efficient protein production systems. Biotechnol Genet Eng Rev 6: 1-41. doi: 10.1080/02648725.1988.10647844
|
| [50] | Brosius J, Erfle M, Storella J (1985) Spacing of the -10 and -35 regions in the tac promoter. Effect on its in vivo activity. J Biol Chem 260: 3539-3541. |
| [51] | Neubauer P, Winter J (2001) Expression and fermentation strategies for recombinant protein production in Escherichia coli. In: Merten OW, Mattanovich D, Lang C et al., editors. Recombinant Protein Production with Prokaryotic and Eukaryotic Cells A Comparative View on Host Physiology. Dordrecht, the Netherlands: Kluwer Academic Publishers; 195-258. |
| [52] |
Craig SP, 3rd, Yuan L, Kuntz DA, et al. (1991) High level expression in Escherichia coli of soluble, enzymatically active schistosomal hypoxanthine/guanine phosphoribosyltransferase and trypanosomal ornithine decarboxylase. Proc Natl Acad Sci U S A 88: 2500-2504. doi: 10.1073/pnas.88.6.2500
|
| [53] |
Elvin CM, Thompson PR, Argall ME, et al. (1990) Modified bacteriophage lambda promoter vectors for overproduction of proteins in Escherichia coli. Gene 87: 123-126. doi: 10.1016/0378-1119(90)90503-J
|
| [54] | Valdez-Cruz NA, Caspeta L, Perez NO, et al. (2010) Production of recombinant proteins in E. coli by the heat inducible expression system based on the phage lambda pL and/or pR promoters. Microb Cell Fact 9: 18. |
| [55] |
Skerra A (1994) Use of the tetracycline promoter for the tightly regulated production of a murine antibody fragment in Escherichia coli. Gene 151: 131-135. doi: 10.1016/0378-1119(94)90643-2
|
| [56] | Guzman LM, Belin D, Carson MJ, et al. (1995) Tight regulation, modulation, and high-level expression by vectors containing the arabinose PBAD promoter. J Bacteriol 177: 4121-4130. |
| [57] |
Siegele DA, Hu JC (1997) Gene expression from plasmids containing the araBAD promoter at subsaturating inducer concentrations represents mixed populations. Proc Natl Acad Sci U S A 94: 8168-8172. doi: 10.1073/pnas.94.15.8168
|
| [58] | Gentz R, Bujard H (1985) Promoters recognized by Escherichia coli RNA polymerase selected by function: highly efficient promoters from bacteriophage T5. J Bacteriol 164: 70-77. |
| [59] | Deuschle U, Kammerer W, Gentz R, et al. (1986) Promoters of Escherichia coli: a hierarchy of in vivo strength indicates alternate structures. EMBO J 5: 2987-2994. |
| [60] | Samuelson J (2011) Bacterial Systems. In: Robinson AS, editor. Production of Membrane Proteins. Weinheim, Germany: Wiley-VCH Verlag GmbH & Co. KGaA; 11-35. |
| [61] |
Studier FW, Moffatt BA (1986) Use of bacteriophage T7 RNA polymerase to direct selective high-level expression of cloned genes. J Mol Biol 189: 113-130. doi: 10.1016/0022-2836(86)90385-2
|
| [62] | Donahue Jr RA, Bebee RL (1999) BL21-SITM competent cells for protein expression in E. coli. Focus 21: 49-51. |
| [63] |
Lowman HB, Bina M (1990) Temperature-mediated regulation and downstream inducible selection for controlling gene expression from the bacteriophage lambda pL promoter. Gene 96: 133-136. doi: 10.1016/0378-1119(90)90353-S
|
| [64] |
Studier FW (1991) Use of bacteriophage T7 lysozyme to improve an inducible T7 expression system. J Mol Biol 219: 37-44. doi: 10.1016/0022-2836(91)90855-Z
|
| [65] |
Lee SK, Keasling JD (2005) A propionate-inducible expression system for enteric bacteria. Appl Environ Microbiol 71: 6856-6862. doi: 10.1128/AEM.71.11.6856-6862.2005
|
| [66] |
Choi YJ, Morel L, Le Francois T, et al. (2010) Novel, versatile, and tightly regulated expression system for Escherichia coli strains. Appl Environ Microbiol 76: 5058-5066. doi: 10.1128/AEM.00413-10
|
| [67] | Saida F, Uzan M, Odaert B, et al. (2006) Expression of highly toxic genes in E. coli: special strategies and genetic tools. Curr Protein Pept Sci 7: 47-56. |
| [68] |
Guan L, Liu Q, Li C, et al. (2013) Development of a Fur-dependent and tightly regulated expression system in Escherichia coli for toxic protein synthesis. BMC Biotechnol 13: 25. doi: 10.1186/1472-6750-13-25
|
| [69] |
Wu H, Pei J, Jiang Y, et al. (2010) pHsh vectors, a novel expression system of Escherichia coli for the large-scale production of recombinant enzymes. Biotechnol Lett 32: 795-801. doi: 10.1007/s10529-010-0223-y
|
| [70] |
Nocadello S, Swennen EF (2012) The new pLAI (lux regulon based auto-inducible) expression system for recombinant protein production in Escherichia coli. Microb Cell Fact 11: 3. doi: 10.1186/1475-2859-11-3
|
| [71] |
Bongers RS, Veening JW, Van Wieringen M, et al. (2005) Development and characterization of a subtilin-regulated expression system in Bacillus subtilis: strict control of gene expression by addition of subtilin. Appl Environ Microbiol 71: 8818-8824. doi: 10.1128/AEM.71.12.8818-8824.2005
|
| [72] |
Ming-Ming Y, Wei-Wei Z, Xi-Feng Z, et al. (2006) Construction and characterization of a novel maltose inducible expression vector in Bacillus subtilis. Biotechnol Lett 28: 1713-1718. doi: 10.1007/s10529-006-9146-z
|
| [73] |
Ming YM, Wei ZW, Lin CY, et al. (2010) Development of a Bacillus subtilis expression system using the improved Pglv promoter. Microb Cell Fact 9: 55. doi: 10.1186/1475-2859-9-55
|
| [74] |
Thuy Le AT, Schumann W (2007) A novel cold-inducible expression system for Bacillus subtilis. Protein Expr Purif 53: 264-269. doi: 10.1016/j.pep.2006.12.023
|
| [75] |
Biedendieck R, Gamer M, Jaensch L, et al. (2007) A sucrose-inducible promoter system for the intra- and extracellular protein production in Bacillus megaterium. J Biotechnol 132: 426-430. doi: 10.1016/j.jbiotec.2007.07.494
|
| [76] |
Gamer M, Frode D, Biedendieck R, et al. (2009) A T7 RNA polymerase-dependent gene expression system for Bacillus megaterium. Appl Microbiol Biotechnol 82: 1195-1203. doi: 10.1007/s00253-009-1952-5
|
| [77] |
Qiu D, Damron FH, Mima T, et al. (2008) PBAD-based shuttle vectors for functional analysis of toxic and highly regulated genes in Pseudomonas and Burkholderia spp. and other bacteria. Appl Environ Microbiol 74: 7422-7426. doi: 10.1128/AEM.01369-08
|
| [78] |
Studier FW (2005) Protein production by auto-induction in high density shaking cultures. Protein Expr Purif 41: 207-234. doi: 10.1016/j.pep.2005.01.016
|
| [79] |
Studier FW (2014) Stable expression clones and auto-induction for protein production in E. coli. Methods Mol Biol 1091: 17-32. doi: 10.1007/978-1-62703-691-7_2
|
| [80] |
Katzen F, Chang G, Kudlicki W (2005) The past, present and future of cell-free protein synthesis. Trends Biotechnol 23: 150-156. doi: 10.1016/j.tibtech.2005.01.003
|
| [81] |
Schwarz D, Dotsch V, Bernhard F (2008) Production of membrane proteins using cell-free expression systems. Proteomics 8: 3933-3946. doi: 10.1002/pmic.200800171
|
| [82] |
Carlson ED, Gan R, Hodgman CE, et al. (2012) Cell-free protein synthesis: applications come of age. Biotechnol Adv 30: 1185-1194. doi: 10.1016/j.biotechadv.2011.09.016
|
| [83] |
Guillerez J, Lopez PJ, Proux F, et al. (2005) A mutation in T7 RNA polymerase that facilitates promoter clearance. Proc Natl Acad Sci U S A 102: 5958-5963. doi: 10.1073/pnas.0407141102
|
| [84] |
Yadav VG, De Mey M, Lim CG, et al. (2012) The future of metabolic engineering and synthetic biology: towards a systematic practice. Metab Eng 14: 233-241. doi: 10.1016/j.ymben.2012.02.001
|
| [85] |
Mijakovic I, Petranovic D, Jensen PR (2005) Tunable promoters in systems biology. Curr Opin Biotechnol 16: 329-335. doi: 10.1016/j.copbio.2005.04.003
|
| [86] |
Pitera DJ, Paddon CJ, Newman JD, et al. (2007) Balancing a heterologous mevalonate pathway for improved isoprenoid production in Escherichia coli. Metab Eng 9: 193-207. doi: 10.1016/j.ymben.2006.11.002
|
| [87] |
Anthony JR, Anthony LC, Nowroozi F, et al. (2009) Optimization of the mevalonate-based isoprenoid biosynthetic pathway in Escherichia coli for production of the anti-malarial drug precursor amorpha-4,11-diene. Metab Eng 11: 13-19. doi: 10.1016/j.ymben.2008.07.007
|
| [88] |
Ajikumar PK, Xiao WH, Tyo KE, et al. (2010) Isoprenoid pathway optimization for Taxol precursor overproduction in Escherichia coli. Science 330: 70-74. doi: 10.1126/science.1191652
|
| [89] |
Elowitz MB, Leibler S (2000) A synthetic oscillatory network of transcriptional regulators. Nature 403: 335-338. doi: 10.1038/35002125
|
| [90] |
Gardner TS, Cantor CR, Collins JJ (2000) Construction of a genetic toggle switch in Escherichia coli. Nature 403: 339-342. doi: 10.1038/35002131
|
| [91] |
Suel GM, Garcia-Ojalvo J, Liberman LM, et al. (2006) An excitable gene regulatory circuit induces transient cellular differentiation. Nature 440: 545-550. doi: 10.1038/nature04588
|
| [92] |
Shetty RP, Endy D, Knight TF, Jr. (2008) Engineering BioBrick vectors from BioBrick parts. J Biol Eng 2: 5. doi: 10.1186/1754-1611-2-5
|
| [93] |
Radeck J, Kraft K, Bartels J, et al. (2013) The Bacillus BioBrick Box: generation and evaluation of essential genetic building blocks for standardized work with Bacillus subtilis. J Biol Eng 7: 29. doi: 10.1186/1754-1611-7-29
|
| [94] |
Hershberg R, Bejerano G, Santos-Zavaleta A, et al. (2001) PromEC: An updated database of Escherichia coli mRNA promoters with experimentally identified transcriptional start sites. Nucleic Acids Res 29: 277. doi: 10.1093/nar/29.1.277
|
| [95] |
Ishii T, Yoshida K, Terai G, et al. (2001) DBTBS: a database of Bacillus subtilis promoters and transcription factors. Nucleic Acids Res 29: 278-280. doi: 10.1093/nar/29.1.278
|
| [96] |
Makita Y, Nakao M, Ogasawara N, et al. (2004) DBTBS: database of transcriptional regulation in Bacillus subtilis and its contribution to comparative genomics. Nucleic Acids Res 32: D75-77. doi: 10.1093/nar/gkh074
|
| [97] |
Sierro N, Makita Y, de Hoon M, et al. (2008) DBTBS: a database of transcriptional regulation in Bacillus subtilis containing upstream intergenic conservation information. Nucleic Acids Res 36: D93-96. doi: 10.1093/nar/gkn421
|
| [98] |
Yao AI, Fenton TA, Owsley K, et al. (2013) Promoter element arising from the fusion of standard BioBrick parts. ACS Synth Biol 2: 111-120. doi: 10.1021/sb300114d
|
| [99] |
de Jong A, Pietersma H, Cordes M, et al. (2012) PePPER: a webserver for prediction of prokaryote promoter elements and regulons. BMC Genomics 13: 299. doi: 10.1186/1471-2164-13-299
|
| [100] |
Klucar L, Stano M, Hajduk M (2010) phiSITE: database of gene regulation in bacteriophages. Nucleic Acids Res 38: D366-370. doi: 10.1093/nar/gkp911
|
| [101] |
Chakravarty A, Carlson JM, Khetani RS, et al. (2007) A novel ensemble learning method for de novo computational identification of DNA binding sites. BMC Bioinformatics 8: 249. doi: 10.1186/1471-2105-8-249
|
| [102] |
Carlson JM, Chakravarty A, DeZiel CE, et al. (2007) SCOPE: a web server for practical de novo motif discovery. Nucleic Acids Res 35: W259-264. doi: 10.1093/nar/gkm310
|
| [103] |
Czar MJ, Cai Y, Peccoud J (2009) Writing DNA with GenoCAD. Nucleic Acids Res 37: W40-47. doi: 10.1093/nar/gkp361
|
| [104] |
Chandran D, Bergmann FT, Sauro HM (2009) TinkerCell: modular CAD tool for synthetic biology. J Biol Eng 3: 19. doi: 10.1186/1754-1611-3-19
|
| [105] |
Hill AD, Tomshine JR, Weeding EM, et al. (2008) SynBioSS: the synthetic biology modeling suite. Bioinformatics 24: 2551-2553. doi: 10.1093/bioinformatics/btn468
|
| [106] |
Kaznessis YN (2011) SynBioSS-aided design of synthetic biological constructs. Methods Enzymol 498: 137-152. doi: 10.1016/B978-0-12-385120-8.00006-1
|
| [107] | Andrianantoandro E, Basu S, Karig DK, et al. (2006) Synthetic biology: new engineering rules for an emerging discipline. Mol Syst Biol2: 2006-0028. |
| [108] |
Endy D (2005) Foundations for engineering biology. Nature 438: 449-453. doi: 10.1038/nature04342
|
| [109] |
Kelly JR, Rubin AJ, Davis JH, et al. (2009) Measuring the activity of BioBrick promoters using an in vivo reference standard. J Biol Eng 3: 4. doi: 10.1186/1754-1611-3-4
|
| [110] |
Carrier TA, Keasling JD (1997) Controlling messenger RNA stability in bacteria: strategies for engineering gene expression. Biotechnol Prog 13: 699-708. doi: 10.1021/bp970095h
|
| [111] |
Carrier TA, Keasling JD (1999) Library of synthetic 5' secondary structures to manipulate mRNA stability in Escherichia coli. Biotechnol Prog 15: 58-64. doi: 10.1021/bp9801143
|
| [112] |
Smolke CD, Carrier TA, Keasling JD (2000) Coordinated, differential expression of two genes through directed mRNA cleavage and stabilization by secondary structures. Appl Environ Microbiol 66: 5399-5405. doi: 10.1128/AEM.66.12.5399-5405.2000
|
| [113] | Gao X, Yuan XX, Shi ZY, et al. (2012) Production of copolyesters of 3-hydroxybutyrate and medium-chain-length 3-hydroxyalkanoates by E. coli containing an optimized PHA synthase gene. Microb Cell Fact 11: 130. |
| [114] |
Salis HM, Mirsky EA, Voigt CA (2009) Automated design of synthetic ribosome binding sites to control protein expression. Nat Biotechnol 27: 946-950. doi: 10.1038/nbt.1568
|
| [115] |
Pfleger BF, Pitera DJ, Smolke CD, et al. (2006) Combinatorial engineering of intergenic regions in operons tunes expression of multiple genes. Nat Biotechnol 24: 1027-1032. doi: 10.1038/nbt1226
|
| [116] |
Dueber JE, Wu GC, Malmirchegini GR, et al. (2009) Synthetic protein scaffolds provide modular control over metabolic flux. Nat Biotechnol 27: 753-759. doi: 10.1038/nbt.1557
|
| [117] |
Topp S, Reynoso CM, Seeliger JC, et al. (2010) Synthetic riboswitches that induce gene expression in diverse bacterial species. Appl Environ Microbiol 76: 7881-7884. doi: 10.1128/AEM.01537-10
|
| [118] |
Wang HH, Isaacs FJ, Carr PA, et al. (2009) Programming cells by multiplex genome engineering and accelerated evolution. Nature 460: 894-898. doi: 10.1038/nature08187
|
| [119] |
Warner JR, Reeder PJ, Karimpour-Fard A, et al. (2010) Rapid profiling of a microbial genome using mixtures of barcoded oligonucleotides. Nat Biotechnol 28: 856-862. doi: 10.1038/nbt.1653
|
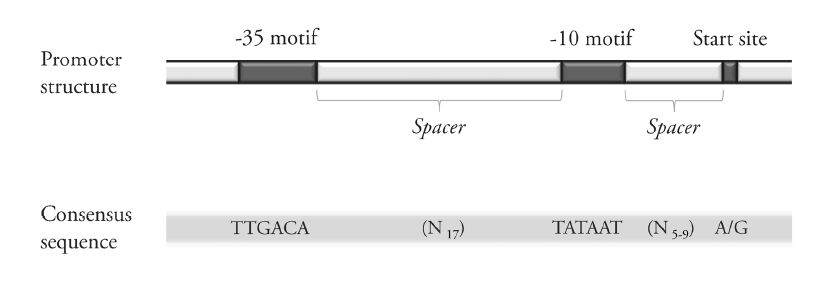
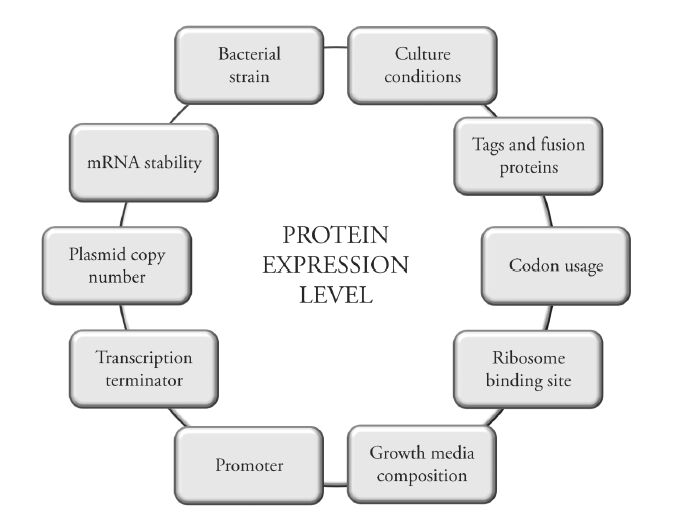
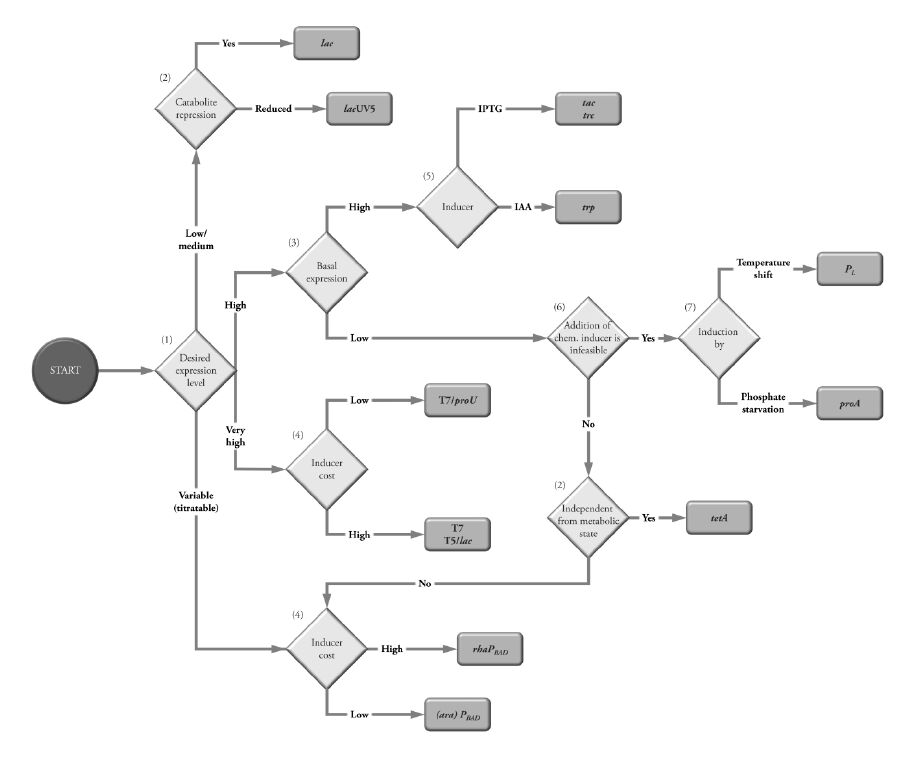
Figures(3) / Tables(3)

Pawel Jajesniak, Tuck Seng Wong. From genetic circuits to industrial-scale biomanufacturing: bacterial promoters as a cornerstone of biotechnology[J]. AIMS Bioengineering, 2015, 2(3): 277-296. doi: 10.3934/bioeng.2015.3.277







 DownLoad:
DownLoad: