Citation: Antonio N. Bojanic, Alejandro Jordán. Modeling the COVID-19 epidemic in Bolivia[J]. Big Data and Information Analytics, 2020, 5(1): 47-57. doi: 10.3934/bdia.2020004

| [1] | Kermack WO and McKendrick AG, (1927) A contribution to the mathematical theory of epidemics. Proc R Soc Lond 115: 700-721. |
| [2] | Lin Q, Zhao S, Gao D, et al. (2020) A conceptual model for the coronavirus disease 2019 (COVID-19) outbreak in Wuhan, China with individual reaction and governmental action. Int J Inf Dis 93: 211-216. |
| [3] | Anastassopoulou C, Russo L, Tsakris A, et al. (2020) Data-based analysis, modelling and forecasting of the COVID-19 outbreak. PloS One 15: e0230405. |
| [4] | Casella F, (2020) Can the COVID-19 epidemic be controlled on the basis of daily test reports?, Preprint, arXiv: 2003.06967. |
| [5] | Giordano G, Blanchini F, Bruno R, et al. (2020) Modelling the COVID-19 epidemic and implementation of population-wide interventions in Italy. Nat Med 26: 855-860. |
| [6] | Wu JT, Leung K, Bushman M, et al. (2020) Estimating clinical severity of COVID-19 from the transmission dynamics in Wuhan, China. Nat Med 26: 506-510. |
| [7] | Tam KM, Walker N and Moreno J, (2020) Projected development of COVID-19 in Louisiana. arXiv 2004.02859. |
| [8] | Hellewell J, Abbott S, Gimma A, et al. (2020) Feasibility of controlling COVID-19 outbreaks by isolation of cases and contacts. Lancet Global Health, 8: e488-e496. |
| [9] | Kucharski AJ, Russell TW, Diamond C, et al. (2020) Early dynamics of transmission and control of COVID-19: a mathematical modelling study, Lancet Infect Dis 20: 553-558. |
| [10] | Birbuet JC and López V R, (2020) Dinámica de expansión del COVID-19 en Bolivia durante las 6 primeras semanas. Working Paper. Available from: https://github.com/rafalopezv/modelo_seir_bolivia. |
| [11] | Tassier T, (2013) The Economics of Epidemiology, Springer-Verlag Berlin Heidelberg. |
| [12] | Munayco CV, Tariq A, Rothenberg R, et al. (2020) Early transmission dynamics of COVID-19 in a southern hemisphere setting: Lima-Peru: February 29th-March 30th, 2020. Infect Dis Model, 5: 338-345. |
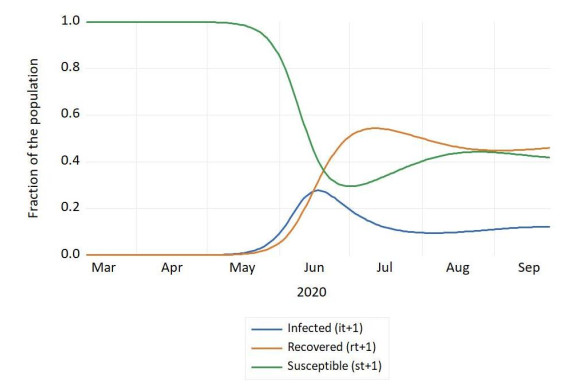
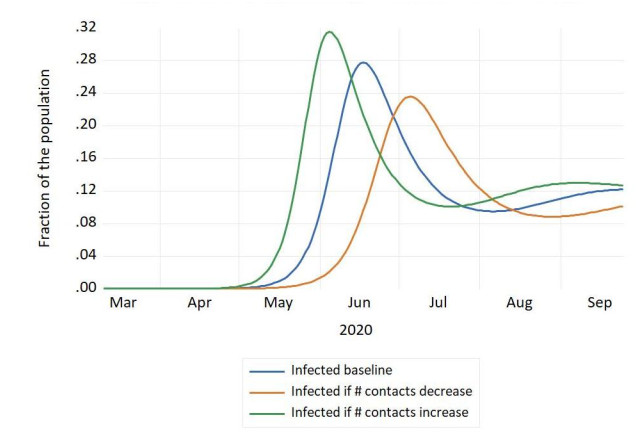
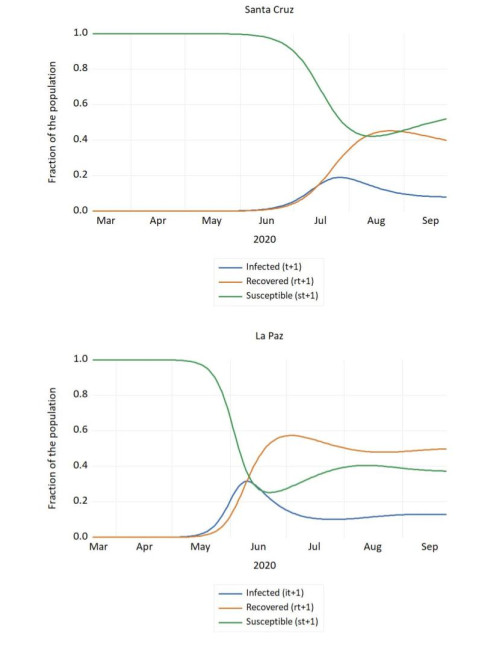
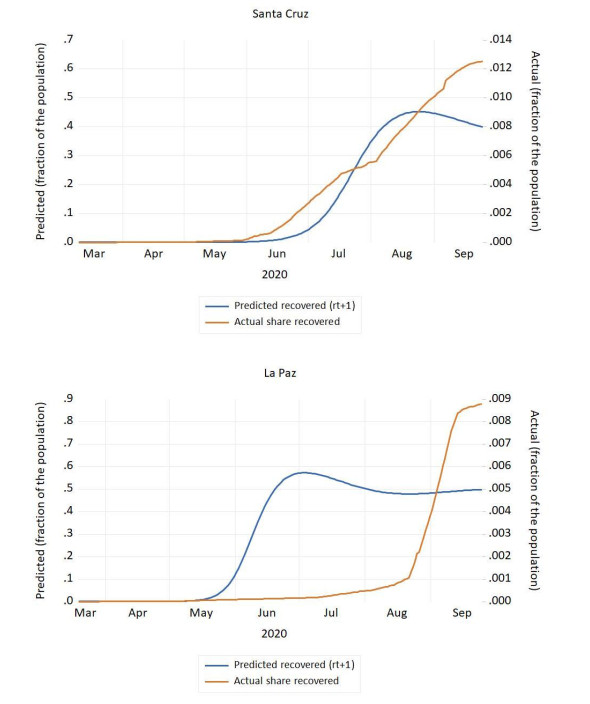
Figures(4) / Tables(1)

Antonio N. Bojanic, Alejandro Jordán. Modeling the COVID-19 epidemic in Bolivia[J]. Big Data and Information Analytics, 2020, 5(1): 47-57. doi: 10.3934/bdia.2020004









 DownLoad:
DownLoad: