Citation: Hannah P Priyanka, Rahul S Nair. Neuroimmunomodulation by estrogen in health and disease[J]. AIMS Neuroscience, 2020, 7(4): 401-417. doi: 10.3934/Neuroscience.2020025

| [1] |
Meites J (1988) Neuroendocrine biomarkers of aging in the rat. Exp Gerontol 23: 349-358. doi: 10.1016/0531-5565(88)90037-X

|
| [2] | Bellinger DL, Lorton D, Lubahn C, et al. (2001) Innervation of lymphoid organs—Association of nerves with cells of the immune system and their implications in disease. Psychoneuroimmunology San Diego: Academic Press, 5-111. |
| [3] |
Downs JL, Wise PM (2009) The role of the brain in female reproductive aging. Mol Cell Endocrinol 299: 32-38. doi: 10.1016/j.mce.2008.11.012
|
| [4] |
Banks WA (2015) The blood-brain barrier in neuroimmunology: tales of separation and assimilation. Brain Behav Immun 44: 1-8. doi: 10.1016/j.bbi.2014.08.007
|
| [5] |
Bellinger DL, Millar BA, Perez S, et al. (2008) Sympathetic modulation of immunity: relevance to disease. Cell Immunol 252: 27-56. doi: 10.1016/j.cellimm.2007.09.005
|
| [6] | ThyagaRajan S, Priyanka HP (2012) Bidirectional communication between the neuroendocrine system and the immune system: relevance to health and diseases. Ann Neurosci 19: 40-46. |
| [7] | Pratap U, Hima L, Kannan T, et al. (2020) Sex-Based Differences in the Cytokine Production and Intracellular Signaling Pathways in Patients With Rheumatoid Arthritis. Arch Rheumatol 35: i-xiii. |
| [8] |
Hima L, Patel MN, Kannan T, et al. (2020) Age-associated decline in neural, endocrine, and immune responses in men and women: Involvement of intracellular signaling pathways. J Neuroimmunol 345: 577290. doi: 10.1016/j.jneuroim.2020.577290
|
| [9] |
ThyagaRajan S, Hima L, Pratap UP, et al. (2019) Estrogen-induced neuroimmunomodulation as facilitator of and barrier to reproductive aging in brain and lymphoid organs. J Chem Neuroanat 95: 6-12. doi: 10.1016/j.jchemneu.2018.02.008
|
| [10] |
Randolph JF, Sowers M, Bondarenko IV, et al. (2004) Change in estradiol and follicle-stimulating hormone across the early menopausal transition: effects of ethnicity and age. J Clin Endocrinol Metab 89: 1555-1561. doi: 10.1210/jc.2003-031183
|
| [11] |
Kermath BA, Gore AC (2012) Neuroendocrine control of the transition to reproductive senescence: lessons learned from the female rodent model. Neuroendocrinol 96: 1-12. doi: 10.1159/000335994
|
| [12] |
Harlow SD, Mitchell ES, Crawford S, et al. (2008) The ReSTAGE Collaboration: defining optimal bleeding criteria for onset of early menopausal transition. Fertil Steril 89: 129-40. doi: 10.1016/j.fertnstert.2007.02.015
|
| [13] |
Wang Y, Mishra A, Brinton RD (2020) Transitions in metabolic and immune systems from pre-menopause to post-menopause: implications for age-associated neurodegenerative diseases. F1000 Res 9: 68. doi: 10.12688/f1000research.21599.1
|
| [14] |
Finkelstein JS, Brockwell SE, Mehta V, et al. (2008) Bone mineral density changes during the menopause transition in a multiethnic cohort of women. J Clin Endocrinol Metab 93: 861-868. doi: 10.1210/jc.2007-1876
|
| [15] |
Brann DW, Mahesh VB (2005) The aging reproductive neuroendocrine axis. Steroids 70: 273-283. doi: 10.1016/j.steroids.2004.12.008
|
| [16] |
Wise PM (2005) Estrogens and cerebrovascular stroke: what do animal models teach us? Ann N Y Acad Sci 1052: 225-232. doi: 10.1196/annals.1347.017
|
| [17] |
Wise PM, Scarbrough K, Lloyd J, et al. (1994) Neuroendocrine concomitants of reproductive aging. Exp Gerontol 29: 275-283. doi: 10.1016/0531-5565(94)90007-8
|
| [18] |
MohanKumar PS, ThyagaRajan S, Quadri SK (1995) Cyclic and age-related changes in norepinephrine concentrations in the medial preoptic area and arcuate nucleus. Brain Res Bull 38: 561-564. doi: 10.1016/0361-9230(95)02031-4
|
| [19] |
MohanKumar PS, ThyagaRajan S, Quadri SK (1997) Tyrosine hydroxylase and DOPA decarboxylase activities in the medial preoptic area and arcuate nucleus during the estrus cycle: effects of aging. Brain Res Bull 42: 265-271. doi: 10.1016/S0361-9230(96)00210-9
|
| [20] |
Wise PM (1982) Norepinephrine and dopamine activity in micro dissected brain areas of the middle-aged and young rat on proestrus. Biol Reprod 27: 562-574. doi: 10.1095/biolreprod27.3.562
|
| [21] |
Wise PM (1984) Estradiol-induced daily luteinizing hormone and prolactin surges in young and middle-aged rats: correlations with age-related changes in pituitary responsiveness and catecholamine turnover rates in micro dissected brain areas. Endocrinology 115: 801-809. doi: 10.1210/endo-115-2-801
|
| [22] |
ThyagaRajan S, Madden KS, Teruya B, et al. (2011) Age-associated alterations in sympathetic noradrenergic innervation of primary and secondary lymphoid organs in female Fischer 344 rats. J Neuroimmunol 233: 54-64. doi: 10.1016/j.jneuroim.2010.11.012
|
| [23] |
Chakrabarti M, Haque A, Banik NL, et al. (2014) Estrogen receptor agonists for attenuation of neuroinflammation and neurodegeneration. Brain Res Bull 109: 22-31. doi: 10.1016/j.brainresbull.2014.09.004
|
| [24] |
Ravichandran KA, Karrunanithi S, Hima L, et al. (2017) Estrogen differentially regulates the expression of tyrosine hydroxylase and nerve growth factor through free radical generation in the thymus and mesenteric lymph nodes of middle-aged ovariectomized female Sprague-Dawley rats. Clin Exp Neuroimmunol 8: 341-350. doi: 10.1111/cen3.12415
|
| [25] |
Priyanka HP, Sharma U, Gopinath S, et al. (2013) Menstrual cycle and reproductive aging alters immune reactivity, NGF expression, antioxidant enzyme activities, and intracellular signalling pathways in the peripheral blood mononuclear cells of healthy women. Brain Behav Immun 32: 131-143. doi: 10.1016/j.bbi.2013.03.008
|
| [26] |
Wise PM, Smith MJ, Dubal DB, et al. (2002) Neuroendocrine modulation and repercussions of female reproductive aging. Recent Prog Horm Res 57: 235-256. doi: 10.1210/rp.57.1.235
|
| [27] |
Murray RW (2001) Estrogen, prolactin, and autoimmunity: actions and interactions. Int Immunopharmacol 1: 995-1008. doi: 10.1016/S1567-5769(01)00045-5
|
| [28] |
Salem ML (2004) Estrogen, a double-edged sword: modulation of TH1- and TH2-mediated inflammations by differential regulation of TH1/TH2 cytokine production. Curr Drug Targets Inflamm Allergy 3: 97-104. doi: 10.2174/1568010043483944
|
| [29] |
Lang TJ (2004) Estrogen as an immunomodulator. Clin Immunol 113: 224-230. doi: 10.1016/j.clim.2004.05.011
|
| [30] | Krzych U, Strausser HR, Bressler JP, et al. (1978) Quantitative differences in immune responses during the various stages of the estrus cycle in female BALB/c mice. J Immunol 121: 1603-1605. |
| [31] |
Straub RH (2007) The complex role of estrogens in inflammation. Endocr Rev 28: 521-574. doi: 10.1210/er.2007-0001
|
| [32] |
Lasarte S, Elsner D, Sanchez-Elsner T, et al. (2013) Estradiol down regulates NF-κB translocation by Ikbkg transcriptional repression in dendritic cells. Genes Immun 14: 462-469. doi: 10.1038/gene.2013.35
|
| [33] |
Maret A, Coudert JD, Garidou L, et al. (2003) Estradiol enhances primary antigen-specific CD4 T cell responses and Th1 development in vivo. Essential role of estrogen receptor alpha expression in hematopoietic cells. Eur J Immunol 33: 512-521. doi: 10.1002/immu.200310027
|
| [34] |
Priyanka HP, Krishnan HC, Singh RV, et al. (2013) Estrogen modulates in vitro T cell responses in a concentration- and receptor-dependent manner: effects on intracellular molecular targets and antioxidant enzymes. Mol Immunol 56: 328-339. doi: 10.1016/j.molimm.2013.05.226
|
| [35] |
Maglione A, Rolla S, Mercanti SF, et al. (2019) The Adaptive Immune System in Multiple Sclerosis: An Estrogen-Mediated Point of View. Cells 8: 1280. doi: 10.3390/cells8101280
|
| [36] |
Ysrraelit MC, Correale J (2019) Impact of sex hormones on immune function and multiple sclerosis development. Immunology 156: 9-22. doi: 10.1111/imm.13004
|
| [37] |
Cutolo M, Sulli A, Capellino S, et al. (2004) Sex hormones influence on the immune system: basic and clinical aspects in autoimmunity. Lupus 13: 635-638. doi: 10.1191/0961203304lu1094oa
|
| [38] |
Chakrabarti M, Haque A, Banik NL, et al. (2014) Estrogen receptor agonists for attenuation of neuroinflammation and neurodegeneration. Brain Res Bull 109: 22-31. doi: 10.1016/j.brainresbull.2014.09.004
|
| [39] |
Babayan AH, Kramár EA (2013) Rapid effects of oestrogen on synaptic plasticity: interactions with actin and its signalling proteins. J Neuroendocrinol 25: 1163-1172. doi: 10.1111/jne.12108
|
| [40] |
Mónica Brauer M, Smith PG (2015) Estrogen and female reproductive tract innervation: cellular and molecular mechanisms of autonomic neuroplasticity. Auton Neurosci 187: 1-17. doi: 10.1016/j.autneu.2014.11.009
|
| [41] |
Priyanka HP, ThyagaRajan S (2013) Selective modulation of lymphoproliferation and cytokine production via intracellular signalling targets by α1- and α2-adrenoceptors and estrogen in splenocytes. Int Immunopharmacol 17: 774-784. doi: 10.1016/j.intimp.2013.08.020
|
| [42] |
Priyanka HP, Pratap UP, Singh RV, et al. (2014) Estrogen modulates β2-adrenoceptor-induced cell-mediated and inflammatory immune responses through ER-α involving distinct intracellular signaling pathways, antioxidant enzymes, and nitric oxide. Cell Immunol 292: 1-8. doi: 10.1016/j.cellimm.2014.08.001
|
| [43] |
Scanzano A, Schembri L, Rasini E, et al. (2015) Adrenergic modulation of migration, CD11b and CD18 expression, ROS and interleukin-8 production by human polymorphonuclear leukocytes. Inflamm Res 64: 127-135. doi: 10.1007/s00011-014-0791-8
|
| [44] |
Prey S, Leaute-Labreze C, Pain C, et al. (2014) Mast cells as possible targets of propranolol therapy: an immunohistological study of beta-adrenergic receptors in infantile haemangiomas. Histopathology 65: 436-439. doi: 10.1111/his.12421
|
| [45] |
Du Y, Yan L, Du H, et al. (2012) β1-adrenergic receptor autoantibodies from heart failure patients enhanced TNF-α secretion in RAW264.7 macrophages in a largely PKA-dependent fashion. J Cell Biochem 113: 3218-3228. doi: 10.1002/jcb.24198
|
| [46] |
Yang H, Du RZ, Qiu JP, et al. (2013) Bisoprolol reverses epinephrine-mediated inhibition of cell emigration through increases in the expression of β-arrestin 2 and CCR7 and PI3K phosphorylation, in dendritic cells loaded with cholesterol. Thromb Res 131: 230-237. doi: 10.1016/j.thromres.2012.12.009
|
| [47] |
Markus T, Hansson SR, Cronberg T, et al. (2010) β-Adrenoceptor activation depresses brain inflammation and is neuroprotective in lipopolysaccharide-induced sensitization to oxygen-glucose deprivation in organotypic hippocampal slices. J Neuroinflammation 7: 94. doi: 10.1186/1742-2094-7-94
|
| [48] |
Cunningham M, Gilkeson G (2011) Estrogen receptors in immunity and autoimmunity. Clin Rev Allergy Immunol 40: 66-73. doi: 10.1007/s12016-010-8203-5
|
| [49] |
Geserick C, Meyer HA, Haendler B (2005) The role of DNA response elements as allosteric modulators of steroid receptor function. Mol Cell Endocrinol 236: 1-7. doi: 10.1016/j.mce.2005.03.007
|
| [50] |
Li J, McMurray RW (2006) Effects of estrogen receptor subtype-selective agonists on immune functions in ovariectomized mice. Int Immunopharmacol 6: 1413-1423. doi: 10.1016/j.intimp.2006.04.019
|
| [51] |
Li J, McMurray RW (2010) Effects of cyclic versus sustained estrogen administration on peripheral immune functions in ovariectomized mice. Am J Reprod Immunol 63: 274-281. doi: 10.1111/j.1600-0897.2009.00784.x
|
| [52] |
Delpy L, Douin-Echinard V, Garidou L, et al. (2005) Estrogen enhances susceptibility to experimental autoimmune myasthenia gravis by promoting type 1-polarized immune responses. J Immunol 175: 5050-5057. doi: 10.4049/jimmunol.175.8.5050
|
| [53] |
Murphy AJ, Guyre PM, Pioli PA (2010) Estradiol suppresses NF-kappa B activation through coordinated regulation of let-7a and miR-125b in primary human macrophages. J Immunol 184: 5029-5037. doi: 10.4049/jimmunol.0903463
|
| [54] |
Tiwari-Woodruff S, Voskuhl RR (2009) Neuroprotective and anti-inflammatory effects of estrogen receptor ligand treatment in mice. J Neurol Sci 286: 81-85. doi: 10.1016/j.jns.2009.04.023
|
| [55] |
Hildebrand F, Hubbard WJ, Choudhry MA, et al. (2006) Are the protective effects of 17 beta-estradiol on splenic macrophages and splenocytes after trauma-haemorrhage mediated via estrogen-receptor (ER)-alpha or ER-beta? J Leukoc Biol 79: 1173-1180. doi: 10.1189/jlb.0106029
|
| [56] |
Kawasaki T, Suzuki T, Choudhry MA, et al. (2010) Salutary effects of 17 beta-estradiol on Peyer's patch T cell functions following trauma-haemorrhage. Cytokine 51: 166-172. doi: 10.1016/j.cyto.2010.03.016
|
| [57] |
Suzuki T, Yu HP, Hsieh YC, et al. (2008) Mitogen activated protein kinase (MAPK) mediates non-genomic pathway of estrogen on T cell cytokine production following trauma-haemorrhage. Cytokine 42: 32-38. doi: 10.1016/j.cyto.2008.02.002
|
| [58] |
Liao ZH, Huang T, Xiao JW, et al. (2019) Estrogen signaling effects on muscle-specific immune responses through controlling the recruitment and function of macrophages and T cells. Skeletal Muscle 9: 20. doi: 10.1186/s13395-019-0205-2
|
| [59] | Khan D, Ansar Ahmed S (2015) The Immune System Is a Natural Target for Estrogen Action: Opposing Effects of Estrogen in Two Prototypical Autoimmune Diseases. Front Immunol 6: 635. |
| [60] | Spengler RN, Allen RM, Remick DG, et al. (1990) Stimulation of alpha-adrenergic receptor augments the production of macrophage-derived tumour necrosis factor. J Immunol 145: 1430-1434. |
| [61] |
Prossnitz ER, Barton M (2011) The G-protein-coupled estrogen receptor GPER in health and disease. Nat Rev Endocrinol 7: 715-726. doi: 10.1038/nrendo.2011.122
|
| [62] |
Bourque M, Dluzen DE, Di Paolo T (2012) Signalling pathways mediating the neuroprotective effects of sex steroids and SERMs in Parkinson's disease. Front Neuroendocrinol 33: 169-178. doi: 10.1016/j.yfrne.2012.02.003
|
| [63] |
Lebesgue D, Chevaleyre V, Zukin RS, et al. (2009) Estradiol rescues neurons from global ischemia-induced cell death: multiple cellular pathways of neuroprotection. Steroids 74: 555-561. doi: 10.1016/j.steroids.2009.01.003
|
| [64] |
Thomas W, Coen N, Faherty S, et al. (2006) Estrogen induces phospholipase A2 activation through ERK1/2 to mobilize intracellular calcium in MCF-7 cells. Steroids 71: 256-265. doi: 10.1016/j.steroids.2005.10.010
|
| [65] |
Titolo D, Mayer CM, Dhillon SS, et al. (2008) Estrogen facilitates both phosphatidylinositol 3-kinase/Akt and ERK1/2 mitogen-activated protein kinase membrane signalling required for long-term neuropeptide Y transcriptional regulation in clonal, immortalized neurons. J Neurosci 28: 6473-6482. doi: 10.1523/JNEUROSCI.0514-08.2008
|
| [66] |
Milette S, Hashimoto M, Perrino S, et al. (2019) Sexual dimorphism and the role of estrogen in the immune microenvironment of liver metastases. Nat Commun 10: 5745. doi: 10.1038/s41467-019-13571-x
|
| [67] |
Wade CB, Dorsa DM (2003) Estrogen activation of cyclic adenosine 5′-monophosphate response element mediated transcription requires the extracellularly regulated kinase/mitogen-activated protein kinase pathway. Endocrinology 144: 832-838. doi: 10.1210/en.2002-220899
|
| [68] |
Fernandez SM, Lewis MC, Pechenino AS, et al. (2008) Estradiol-induced enhancement of object memory consolidation involves hippocampal ERK activation and membrane-bound estrogen receptors. J Neurosci 28: 8660-8667. doi: 10.1523/JNEUROSCI.1968-08.2008
|
| [69] |
Carlstrom L, Ke ZJ, Unnerstall JR, et al. (2001) Estrogen modulation of the cyclic AMP response element-binding protein pathway. Effects of long-term and acute treatments. Neuroendocrinology 74: 227-243. doi: 10.1159/000054690
|
| [70] |
Grove-Strawser D, Boulware MI, Mermelstein PG (2010) Membrane estrogen receptors activate the metabotropic glutamate receptors mGluR5 and mGluR3 to bidirectionally regulate CREB phosphorylation in female rat striatal neurons. Neuroscience 170: 1045-1055. doi: 10.1016/j.neuroscience.2010.08.012
|
| [71] |
Sanchez MG, Morissette M, Di Paolo T (2012) Effect of a chronic treatment with 17 β-estradiol on striatal dopamine neurotransmission and the Akt/GSK3 signalling pathway in the brain of ovariectomized monkeys. Psychoneuroendocrinology 37: 280-291. doi: 10.1016/j.psyneuen.2011.06.012
|
| [72] |
Zhang QG, Wang R, Tang H, et al. (2014) Brain-derived estrogen exerts anti-inflammatory and neuroprotective actions in the rat hippocampus. Mol Cell Endocrinol 389: 84-91. doi: 10.1016/j.mce.2013.12.019
|
| [73] |
Pratap UP, Patil A, Sharma HR, et al. (2016) Estrogen-induced neuroprotective and anti-inflammatory effects are dependent on the brain areas of middle-aged female rats. Brain Res Bull 124: 238-253. doi: 10.1016/j.brainresbull.2016.05.015
|
| [74] |
Maharjan S, Serova LI, Sabban EL (2010) Membrane-initiated estradiol signalling increases tyrosine hydroxylase promoter activity with ER alpha in PC12 cells. J Neurochem 112: 42-55. doi: 10.1111/j.1471-4159.2009.06430.x
|
| [75] |
Pendergast JS, Tuesta LM, Bethea JR (2008) Oestrogen receptor beta contributes to the transient sex difference in tyrosine hydroxylase expression in the mouse locus coeruleus. J Neuroendocrinol 20: 1155-1164. doi: 10.1111/j.1365-2826.2008.01776.x
|
| [76] |
Thanky NR, Son JH, Herbison AE (2002) Sex differences in the regulation of tyrosine hydroxylase gene transcription by estrogen in the locus coeruleus of TH9-LacZ transgenic mice. Brain Res Mol Brain Res 104: 220-226. doi: 10.1016/S0169-328X(02)00383-2
|
| [77] |
Yamaguchi N, Yuri K (2014) Estrogen-dependent changes in estrogen receptor-β mRNA expression in middle-aged female rat brain. Brain Res 1543: 49-57. doi: 10.1016/j.brainres.2013.11.010
|
| [78] |
Foster TC (2012) Role of estrogen receptor alpha and beta expression and signalling on cognitive function during aging. Hippocampus 22: 656-669. doi: 10.1002/hipo.20935
|
| [79] | Coyoy-Salgado A, Segura-Uribe JJ, Manuel Gallardo J, et al. (2020) Tibolone regulates systemic metabolism and the expression of sex hormone receptors in the central nervous system of ovariectomised rats fed with high-fat and high-fructose diet. Brain Res 6: 1748. |
| [80] | Zoubina EV, Mize AL, Alper RH, et al. (2001) Acute and chronic estrogen supplementation decreases uterine sympathetic innervation in ovariectomized adult virgin rats. Histol Histopathol 16: 989-996. |
| [81] |
Chisholm NC, Packard AR, Koss WA, et al. (2012) The Effects of Long-Term Treatment with Estradiol and Medroxyprogesterone Acetate on Tyrosine Hydroxylase Fibers and Neuron Number in the Medial Prefrontal Cortex of Aged Female Rats. Endocrinology 153: 4874-4882. doi: 10.1210/en.2012-1412
|
| [82] |
Kale P, Mohanty A, Patil A, et al. (2014) Estrogen modulates neural-immune interactions through intracellular signalling pathways and antioxidant enzyme activity in the spleen of middle-aged ovariectomized female rats. J Neuroimmunol 267: 7-15. doi: 10.1016/j.jneuroim.2013.11.003
|
| [83] |
Tang MX, Jacobs D, Stern Y, et al. (1996) Effect of oestrogen during menopause on risk and age at onset of Alzheimer's disease. Lancet 348: 429-432. doi: 10.1016/S0140-6736(96)03356-9
|
| [84] |
Jacome LF, Gautreaux C, Inagaki T, et al. (2010) Estradiol and ERβ agonists enhance recognition memory, and DPN, an ERβ agonist, alters brain monoamines. Neurobiol Learn Mem 94: 488-498. doi: 10.1016/j.nlm.2010.08.016
|
| [85] |
Gibbs RB (2010) Estrogen therapy and cognition: a review of the cholinergic hypothesis. Endocr Rev 31: 224-253. doi: 10.1210/er.2009-0036
|
| [86] |
Norbury R, Cutter WJ, Compton J, et al. (2003) The neuroprotective effects of estrogen on the aging brain. Exp Gerontol 38: 109-117. doi: 10.1016/S0531-5565(02)00166-3
|
| [87] | Paul V, Ekambaram P (2011) Involvement of nitric oxide in learning & memory processes. Indian J Med Res 133: 471-478. |
| [88] |
Kopf SR, Benton RS, Kalfin R, et al. (2001) NO synthesis inhibition decreases cortical ACH release and impairs retention of conditioned response. Brain Res 894: 141-144. doi: 10.1016/S0006-8993(00)03148-6
|
| [89] |
Ghisletti S, Meda C, Maggi A, et al. (2005) 17 Beta-estradiol inhibits inflammatory gene expression by controlling NF-kappa B intracellular localization. Mol Cell Biol 25: 2957-2968. doi: 10.1128/MCB.25.8.2957-2968.2005
|
| [90] |
Walf AA, Paris JJ, Rhodes ME, et al. (2011) Divergent mechanisms for trophic actions of estrogens in the brain and peripheral tissues. Brain Res 1379: 119-136. doi: 10.1016/j.brainres.2010.11.081
|
| [91] |
Hasan W, Smith HJ, Ting AY, et al. (2005) Estrogen alters trkA and p75 neurotrophin receptor expression within sympathetic neurons. J Neurobiol 65: 192-204. doi: 10.1002/neu.20183
|
| [92] |
Arbogast LA, Hyde JF (2000) Estradiol attenuates the forskolin-induced increase in hypothalamic tyrosine hydroxylase activity. Neuroendocrinology 71: 219-227. doi: 10.1159/000054539
|
| [93] |
Kritzer MF, Kohama SG (1998) Ovarian hormones influence the morphology, distribution, and density of tyrosine hydroxylase immunoreactive axons in the dorsolateral prefrontal cortex of adult rhesus monkeys. J Comp Neurol 395: 1-17. doi: 10.1002/(SICI)1096-9861(19980525)395:1<1::AID-CNE1>3.0.CO;2-4
|
| [94] |
Babu GN, Vijayan E (1984) Hypothalamic tyrosine hydroxylase activity and plasma gonadotropin and prolactin levels in ovariectomized-steroid treated rats. Brain Res Bull 12: 555-558. doi: 10.1016/0361-9230(84)90171-0
|
| [95] |
Chisholm NC, Packard AR, Koss WA, et al. (2012) The effects of long-Term treatment with estradiol and medroxyprogesterone acetate on tyrosine hydroxylase fibers and neuron number in the medial prefrontal cortex of aged female rats. Endocrinology 153: 4874-4882. doi: 10.1210/en.2012-1412
|
| [96] |
Turcano P, Savica R (2020) Sex differences in movement disorders. Handb Clin Neurol 175: 275-282. doi: 10.1016/B978-0-444-64123-6.00019-9
|
| [97] |
Esterbauer H, Schaur RJ, Zollner H (1991) Chemistry and biochemistry of 4-hydroxynonenal, malonaldehyde and related aldehydes. Free Radic Biol Med 11: 81-128. doi: 10.1016/0891-5849(91)90192-6
|
| [98] |
Viveros MP, Arranz L, Hernanz A, et al. (2007) A model of premature aging in mice based on altered stress-related behavioural response and immunosenescence. Neuroimmunomodulation 14: 157-162. doi: 10.1159/000110640
|
| [99] |
Tian L, Cai Q, Bowen R, et al. (1995) Effects of caloric restriction on age-related oxidative modifications of macromolecules and lymphocyte proliferation in rats. Free Radic Biol Med 19: 859-865. doi: 10.1016/0891-5849(95)00090-K
|
| [100] |
Baeza I, Alvarado C, Alvarez P, et al. (2009) Improvement of leucocyte functions in ovariectomised aged rats after treatment with growth hormone, melatonin, oestrogens or phyto-oestrogens. J Reprod Immunol 80: 70-79. doi: 10.1016/j.jri.2009.02.002
|
| [101] |
Baeza I, De Castro NM, Giménez-Llort L, et al. (2010) Ovariectomy, a model of menopause in rodents, causes a premature aging of the nervous and immune systems. J Neuroimmunol 219: 90-99. doi: 10.1016/j.jneuroim.2009.12.008
|
| [102] |
Baeza I, Fdez-Tresguerres J, Ariznavarreta C, et al. (2010) Effects of growth hormone, melatonin, oestrogens and phytoestrogens on the oxidized glutathione (GSSG)/reduced glutathione (GSH) ratio and lipid peroxidation in aged ovariectomized rats. Biogerontology 11: 687-701. doi: 10.1007/s10522-010-9282-7
|
| [103] |
Vina J, Gambini J, Lopez-Grueso R, et al. (2011) Females live longer than males: role of oxidative stress. Curr Pharm Des 17: 3959-3965. doi: 10.2174/138161211798764942
|
| [104] |
Germain D (2016) Sirtuins and the Estrogen Receptor as Regulators of the Mammalian Mitochondrial UPR in Cancer and Aging. Adv Cancer Res 130: 211-256. doi: 10.1016/bs.acr.2016.01.004
|
| [105] |
Brann D, Raz L, Wang R, et al. (2012) Oestrogen signalling and neuroprotection in cerebral ischaemia. J Neuroendocrinol 24: 34-47. doi: 10.1111/j.1365-2826.2011.02185.x
|
| [106] |
Simpkins JW, Green PS, Gridley KE, et al. (1997) Role of estrogen replacement therapy in memory enhancement and the prevention of neuronal loss associated with Alzheimer's disease. Am J Med 103: 19S-25S. doi: 10.1016/S0002-9343(97)00260-X
|
| [107] | Shulman GI, Barrett EJ, Sherwin RS (2003) Integrated fuel metabolism. Ellenberg & Rifkin's diabetes mellitus NewYork: McGraw-Hill, 1-13. |
| [108] | Shi J, Simpkins JW (1997) 17 beta-Estradiol modulation of glucose transporter 1 expression in blood-brain barrier. Am J Physiol 272: E1016-E1022. |
| [109] |
Kostanyan A, Nazaryan A (1992) Rat brain glycolysis regulation by estradiol-17 beta. Biochim Biophys Acta 1133: 301-306. doi: 10.1016/0167-4889(92)90051-C
|
| [110] | Magistretti P (2008) Brain energy metabolism. Fundamental neuroscience San Diego: Academic, 271-296. |
| [111] |
Hernández-R J (1992) Na+/K(+)-ATPase regulation by neurotransmitters. Neurochem Int 20: 1-10. doi: 10.1016/0197-0186(92)90119-C
|
| [112] |
Brinton RD (2008) Estrogen regulation of glucose metabolism and mitochondrial function: therapeutic implications for prevention of Alzheimer's disease. Adv Drug Deliv Rev 60: 1504-1511. doi: 10.1016/j.addr.2008.06.003
|
| [113] |
Kostanyan A, Nazaryan A (1992) Rat brain glycolysis regulation by estradiol-17 beta. Biochim Biophys Acta 1133: 301-306. doi: 10.1016/0167-4889(92)90051-C
|
| [114] |
Moorthy K, Yadav UC, Siddiqui MR, et al. (2004) Effect of estradiol and progesterone treatment on carbohydrate metabolizing enzymes in tissues of aging female rats. Biogerontology 5: 249-259. doi: 10.1023/B:BGEN.0000038026.89337.02
|
| [115] |
Rasgon NL, Silverman D, Siddarth P, et al. (2005) Estrogen use and brain metabolic change in postmenopausal women. Neurobiol Aging 26: 229-235. doi: 10.1016/j.neurobiolaging.2004.03.003
|
| [116] |
Monteiro R, Teixeira D, Calhau C (2014) Estrogen signaling in metabolic inflammation. Mediators Inflamm 2014: 615917. doi: 10.1155/2014/615917
|
| [117] |
Villa A, Rizzi N, Vegeto E, et al. (2015) Estrogen accelerates the resolution of inflammation in macrophagic cells. Sci Rep 5: 15224. doi: 10.1038/srep15224
|
| [118] |
Bagger YZ, Tankó LB, Alexandersen P, et al. (2005) Early postmenopausal hormone therapy may prevent cognitive impairment later in life. Menopause 12: 12-7. doi: 10.1097/00042192-200512010-00005
|
| [119] |
Maki PM (2006) Hormone therapy and cognitive function: is there a critical period for benefit? Neuroscience 138: 1027-1030. doi: 10.1016/j.neuroscience.2006.01.001
|
| [120] |
Keller JN, Germeyer A, Begley JG, et al. (1997) 17 Beta-estradiol attenuates oxidative impairment of synaptic Na+/K+-ATPase activity, glucose transport, and glutamate transport induced by amyloid beta-peptide and iron. J Neurosci Res 50: 522-530. doi: 10.1002/(SICI)1097-4547(19971115)50:4<522::AID-JNR3>3.0.CO;2-G
|
| [121] |
Kumar P, Kale RK, McLean P, et al. (2011) Protective effects of 17 β-estradiol on altered age related neuronal parameters in female rat brain. Neurosci Lett 502: 56-60. doi: 10.1016/j.neulet.2011.07.024
|
| [122] |
Shi C, Xu J (2008) Increased vulnerability of brain to estrogen withdrawal-induced mitochondrial dysfunction with aging. J Bioenerg Biomembr 40: 625-630. doi: 10.1007/s10863-008-9195-1
|
| [123] |
Wong-Riley MT (1989) Cytochrome oxidase: an endogenous metabolic marker for neuronal activity. Trends Neurosci 12: 94-101. doi: 10.1016/0166-2236(89)90165-3
|
| [124] |
Bettini E, Maggi A (1992) Estrogen induction of cytochrome c oxidase subunit III in rat hippocampus. J Neurochem 58: 1923-1929. doi: 10.1111/j.1471-4159.1992.tb10070.x
|
| [125] |
Soane L, Kahraman S, Kristian T, et al. (2007) Mechanisms of impaired mitochondrial energy metabolism in acute and chronic neurodegenerative disorders. J Neurosci Res 85: 3407-3415. doi: 10.1002/jnr.21498
|
| [126] |
Atamna H, Frey WH (2007) Mechanisms of mitochondrial dysfunction and energy deficiency in Alzheimer's disease. Mitochondrion 7: 297-310. doi: 10.1016/j.mito.2007.06.001
|
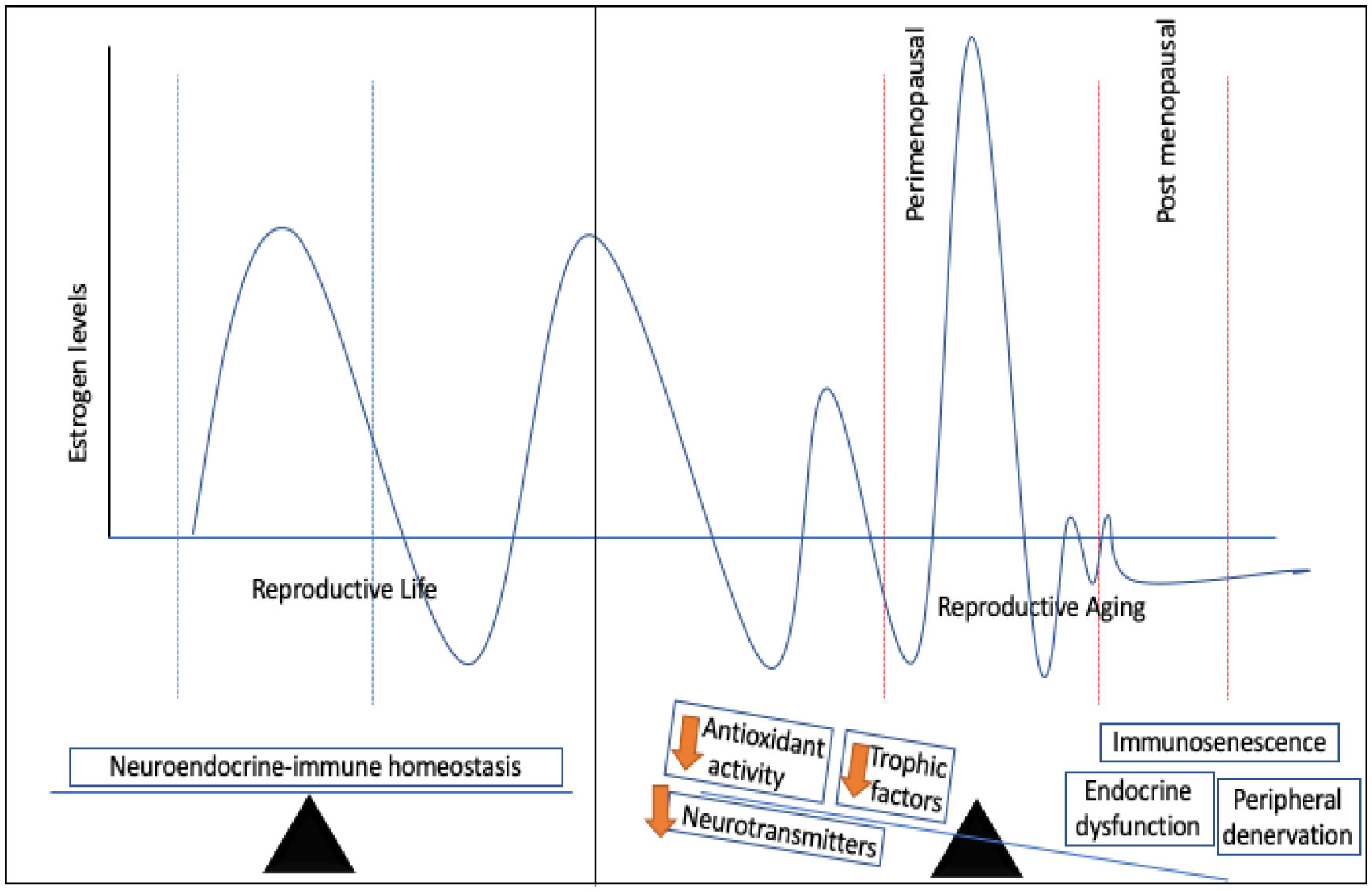
Figures(1)

Hannah P Priyanka, Rahul S Nair. Neuroimmunomodulation by estrogen in health and disease[J]. AIMS Neuroscience, 2020, 7(4): 401-417. doi: 10.3934/Neuroscience.2020025







 DownLoad:
DownLoad: