Citation: Daniele D’Arrigo, Marta Bottagisio, Silvia Lopa, Matteo Moretti, Arianna B. Lovati. Tissue engineering approaches to develop decellularized tendon matrices functionalized with progenitor cells cultured under undifferentiated and tenogenic conditions[J]. AIMS Bioengineering, 2017, 4(4): 431-445. doi: 10.3934/bioeng.2017.4.431

| [1] |
Andia I, Maffulli N (2015) Muscle and tendon injuries: the role of biological interventions to and assist healing and recovery. Arthroscopy 31: 999–1015. doi: 10.1016/j.arthro.2014.11.024

|
| [2] | Bottagisio M, Lovati AB (2017) A review on animal models and treatments for the reconstruction of Achilles and flexor tendons. J Mater Sci Mater Med 28: 1–15. |
| [3] |
Hopkins C, Fu SC, Chua E, et al. (2016) Critical review on the socio-economic impact of tendinopathy. Asia-Pacific J Sport Med Arthrosc Rehabil Technol 4: 9–20. doi: 10.1016/j.asmart.2016.01.002
|
| [4] |
Docheva D, Müller SA, Majewski M, et al. (2015) Biologics for tendon repair. Adv Drug Deliv Rev 84: 222–239. doi: 10.1016/j.addr.2014.11.015
|
| [5] | Petrou IG, Grognuz A, Hirt-Burri N, et al. (2014) Cell therapies for tendons: old cell choice for modern innovation. Swiss Med Wkly 144: 1–11. |
| [6] |
Legrand A, Kaufman Y, Long C, et al. (2017) Molecular biology of flexor tendon healing in relation to reduction of tendon adhesions. J Hand Surg Am 42: 722–726. doi: 10.1016/j.jhsa.2017.06.013
|
| [7] | Akpancar S, Tatar O, Turgut H, et al. (2016) The current perspectives of stem cell therapy in orthopedic surgery. Arch trauma Res 5: e37976. |
| [8] |
Wobma H, Vunjak-Novakovic G (2016) Tissue engineering and regenerative medicine 2015: a year in review. Tissue Eng Part B Rev 22: 101–113. doi: 10.1089/ten.teb.2015.0535
|
| [9] | Lovati AB, Bottagisio M, Moretti M (2016) Decellularized and engineered tendons as biological substitutes: a critical review. Stem Cells Int 2016: 7276150. |
| [10] | Bottagisio M, Pellegata AF, Boschetti F, et al. (2016) A new strategy for the decellularisation of large equine tendons as biocompatible tendon substitutes. Eur Cells Mater 32: 58–73. |
| [11] |
Pellegata AF, Bottagisio M, Boschetti F, et al. (2017) Terminal sterilization of equine-derived decellularized tendons for clinical use. Mater Sci Eng C 75: 43–49. doi: 10.1016/j.msec.2017.02.009
|
| [12] |
Wang S, Wang Y, Song L, et al. (2017) Decellularized tendon as a prospective scaffold for tendon repair. Mater Sci Eng C 77: 1290–1301. doi: 10.1016/j.msec.2017.03.279
|
| [13] |
Yao L, Bestwick CS, Bestwick L, et al. (2006) Phenotypic drift in human tenocyte culture. Tissue Eng 12: 1843–1849. doi: 10.1089/ten.2006.12.1843
|
| [14] |
Schwarz R, Colarusso L, Doty P (1976) Maintenance of differentiation in primary cultures of avian tendon cells. Exp Cell Res 102: 63–71. doi: 10.1016/0014-4827(76)90299-8
|
| [15] |
Van Eijk F, Saris DBF, Riesle J, et al. (2004) Tissue engineering of ligaments: a comparison of bone marrow stromal cells, anterior cruciate ligament, and skin fibroblasts as cell source. Tissue Eng 10: 893–903. doi: 10.1089/1076327041348428
|
| [16] |
Hankemeier S, Hurschler C, Zeichen J, et al. (2009) Bone marrow stromal cells in a liquid fibrin matrix improve the healing process of patellar tendon window defects. Tissue Eng Part A 15: 1019–1030. doi: 10.1089/ten.tea.2008.0046
|
| [17] | Shenaq DS, Rastegar F, Petkovic D, et al. (2010) Mesenchymal progenitor cells and their orthopedic applications: forging a path towards clinical trials. Stem Cells Int 2010: 519028. |
| [18] | Contreras-Kallens P, Terraza C, Oyarce K, et al. (2017) Mesenchymal stem cells and their immunosuppressive role in transplantation tolerance. Ann Ny Acad Sci: 1–22. |
| [19] |
Bottagisio M, Lopa S, Granata V, et al. (2017) Different combinations of growth factors for the tenogenic differentiation of bone marrow mesenchymal stem cells in monolayer culture and in fibrin-based three-dimensional constructs. Differentiation 95: 44–53. doi: 10.1016/j.diff.2017.03.001
|
| [20] |
Goncalves AI, Rodrigues MT, Lee SJ, et al. (2013) Understanding the role of growth factors in modulating stem cell tenogenesis. PLoS One 8: e83734. doi: 10.1371/journal.pone.0083734
|
| [21] | Gaspar D, Spanoudes K, Holladay C, et al. (2014) Progress in cell-based therapies for tendon repair. Adv Drug Deliv Rev 84: 240–256. |
| [22] |
Spanoudes K, Gaspar D, Pandit A, et al. (2014) The biophysical, biochemical, and biological toolbox for tenogenic phenotype maintenance in vitro. Trends Biotechnol 32: 474–482. doi: 10.1016/j.tibtech.2014.06.009
|
| [23] |
Li VC, Kirschner MW (2014) Molecular ties between the cell cycle and differentiation in embryonic stem cells. Proc Natl Acad Sci USA 111: 9503–9508. doi: 10.1073/pnas.1408638111
|
| [24] | Ozasa Y, Amadio PC, Thoreson AR, et al. (2013) Repopulation of intrasynovial flexor tendon allograft with bone marrow stromal cells: an ex vivo model. Tissue Eng Part A 20: 566–574. |
| [25] | Kryger GS, Chong AKS, Costa M, et al. (2007) A comparison of tenocytes and mesenchymal stem cells for use in flexor tendon tissue engineering. J Hand Surgery Am 32: 597–605. |
| [26] |
Thorfinn J, Saber S, Angelidis IK, et al. (2009) Flexor tendon tissue engineering: temporal distribution of donor tenocytes versus recipient cells. Plast Reconstr Surg 124: 2019–2026. doi: 10.1097/PRS.0b013e3181bcf320
|
| [27] |
Taylor SH, Al-Youha S, van Agtmael T, et al. (2011) Tendon is covered by a basement membrane epithelium that is required for cell retention and the prevention of adhesion formation. PLoS One 6: e16337. doi: 10.1371/journal.pone.0016337
|
| [28] |
Angelidis IK, Thorfinn J, Connolly ID, et al. (2010) Tissue engineering of flexor tendons: The effect of a tissue bioreactor on adipoderived stem cellseeded and fibroblast-seeded tendon constructs. J Hand Surg Am 35: 1466–1472. doi: 10.1016/j.jhsa.2010.06.020
|
| [29] |
Qin TW, Sun YL, Thoreson AR, et al. (2015) Effect of mechanical stimulation on bone marrow stromal cell-seeded tendon slice constructs: a potential engineered tendon patch for rotator cuff repair. Biomaterials 51: 43–50. doi: 10.1016/j.biomaterials.2015.01.070
|
| [30] |
Youngstrom DW, Rajpar I, Kaplan DL, et al. (2015) A bioreactor system for in vitro tendon differentiation and tendon tissue engineering. J Orthop Res 33: 911–918. doi: 10.1002/jor.22848
|
| [31] |
Schulze-Tanzil G, Al-Sadi O, Ertel W, et al. (2012) Decellularized tendon extracellular matrix-a valuable approach for tendon reconstruction? Cells 1: 1010–1028. doi: 10.3390/cells1041010
|
| [32] |
Fitzpatrick LE, McDevitt TC (2015) Cell-derived matrices for tissue engineering and regenerative medicine applications. Biomater Sci 3: 12–24. doi: 10.1039/C4BM00246F
|
| [33] | Sharma P, Maffulli N (2006) Biology of tendon injury: healing, modeling and remodeling. J Musculoskelet Neuronal Interact 6: 181–190. |
| [34] |
Herberts CA, Kwa MS, Hermsen HP (2011) Risk factors in the development of stem cell therapy. J Transl Med 9: 29. doi: 10.1186/1479-5876-9-29
|
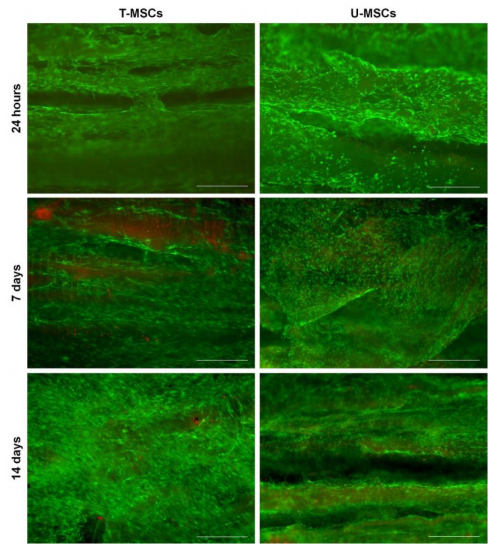
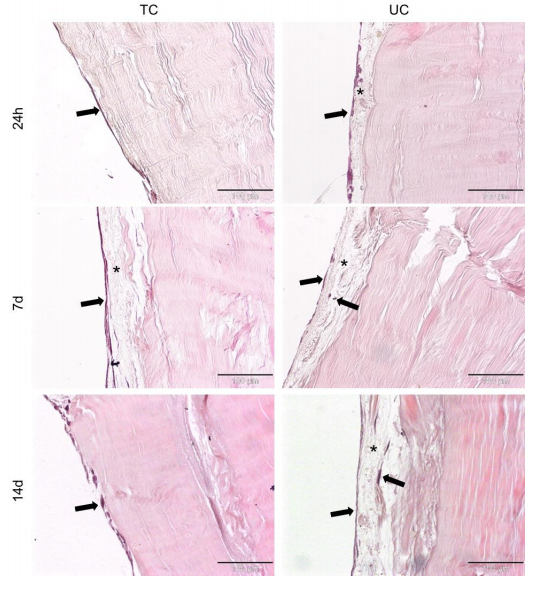
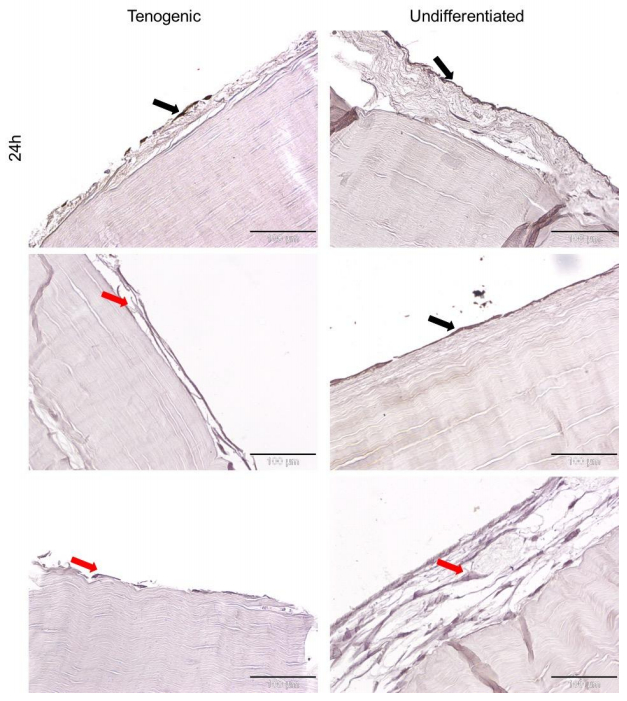
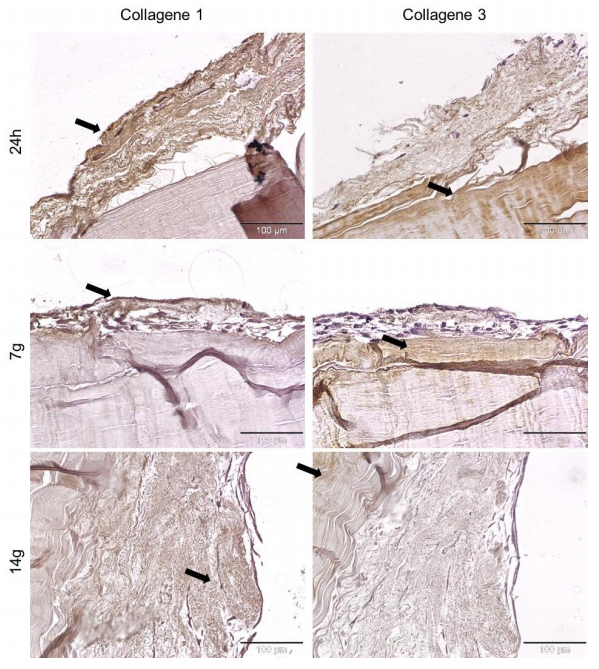
Figures(5)

Daniele D’Arrigo, Marta Bottagisio, Silvia Lopa, Matteo Moretti, Arianna B. Lovati. Tissue engineering approaches to develop decellularized tendon matrices functionalized with progenitor cells cultured under undifferentiated and tenogenic conditions[J]. AIMS Bioengineering, 2017, 4(4): 431-445. doi: 10.3934/bioeng.2017.4.431







 DownLoad:
DownLoad: