Citation: Yihan Zhang. Manufacture of complex heart tissues: technological advancements and future directions[J]. AIMS Bioengineering, 2021, 8(1): 73-92. doi: 10.3934/bioeng.2021008

| [1] | Cardiovascular diseases (CVDs), World Health Organization Available from: https://www.who.int/health-topics/cardiovascular-diseases. |
| [2] |
Fuchs M, Schibilsky D, Zeh W, et al. (2019) Does the heart transplant have a future? Eur J Cardio-thorac 55: i38-i48. doi: 10.1093/ejcts/ezz107

|
| [3] |
Qasim M, Haq F, Kang M, et al. (2019) 3D printing approaches for cardiac tissue engineering and role of immune modulation in tissue regeneration. Int J Nanomed 14: 1311-1333. doi: 10.2147/IJN.S189587
|
| [4] |
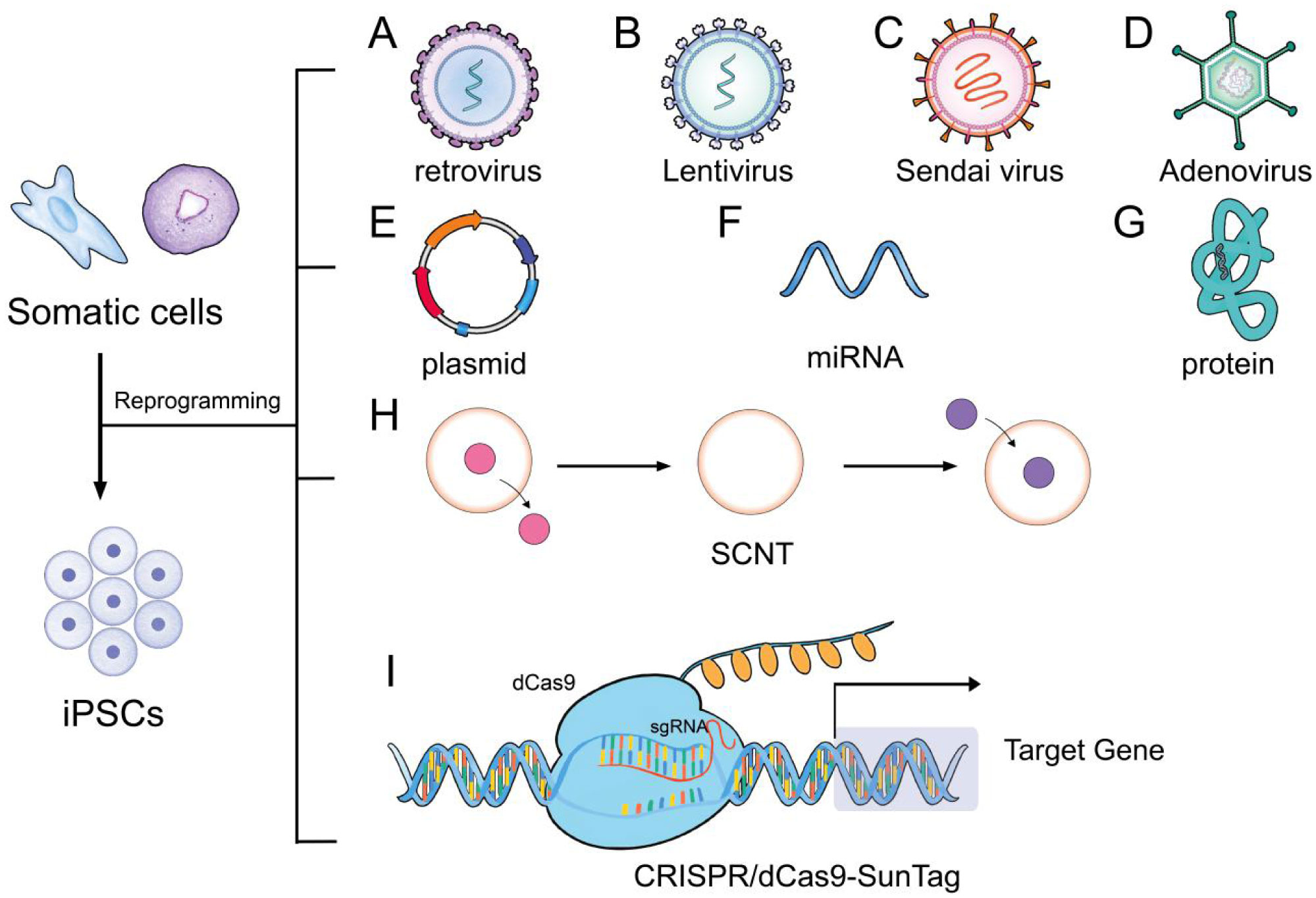
Takahashi K, Yamanaka S (2006) Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 126: 663-676. doi: 10.1016/j.cell.2006.07.024
|
| [5] |
Smith Z, Sindhu C, Meissner A (2016) Molecular features of cellular reprogramming and development. Nat Rev Mol Cell Biol 17: 139-154. doi: 10.1038/nrm.2016.6
|
| [6] |
Okita K, Ichisaka T, Yamanaka S (2007) Generation of germline-competent induced pluripotent stem cells. Nature 448: 313-317. doi: 10.1038/nature05934
|
| [7] |
Yoshida Y, Yamanaka S (2017) Induced pluripotent stem cells 10 years later. Circ Res 120: 1958-1968. doi: 10.1161/CIRCRESAHA.117.311080
|
| [8] |
Maherali N, Sridharan R, Xie W, et al. (2007) Directly reprogrammed fibroblasts show global epigenetic remodeling and widespread tissue contribution. Cell Stem Cell 1: 55-70. doi: 10.1016/j.stem.2007.05.014
|
| [9] |
Somers A, Jean JC, Sommer CA, et al. (2010) Generation of transgene-free lung disease-specific human induced pluripotent stem cells using a single excisable lentiviral stem cell cassette. Stem cells 28: 1728-1740. doi: 10.1002/stem.495
|
| [10] | Fusaki N, Ban H, Nishiyama A, et al. (2009) Efficient induction of transgene-free human pluripotent stem cells using a vector based on Sendai virus, an RNA virus that does not integrate into the host genome. P Jpn Acad B Phys 85: 348-362. |
| [11] |
Carey BW, Markoulaki S, Hanna J, et al. (2009) Reprogramming of murine and human somatic cells using a single polycistronic vector. Proc Natl Acad Sci USA 106: 157-162. doi: 10.1073/pnas.0811426106
|
| [12] |
Stadtfeld M, Nagaya M, Utikal J, et al. (2008) Induced pluripotent stem cells generated without viral integration. Science 322: 945-949. doi: 10.1126/science.1162494
|
| [13] |
McLenachan S, Sarsero J, Ioannou P (2007) Flow-cytometric analysis of mouse embryonic stem cell lipofection using small and large DNA constructs. Genomics 89: 708-720. doi: 10.1016/j.ygeno.2007.02.006
|
| [14] |
Subramanyam D, Lamouille S, Judson R, et al. (2011) Multiple targets of miR-302 and miR-372 promote reprogramming of human fibroblasts to induced pluripotent stem cells. Nat Biotechnol 29: 443-448. doi: 10.1038/nbt.1862
|
| [15] |
Kim D, Kim CH, Moon JI, et al. (2009) Generation of human induced pluripotent stem cells by direct delivery of reprogramming proteins. Cell Stem Cell 4: 472-476. doi: 10.1016/j.stem.2009.05.005
|
| [16] |
Warren L, Manos PD, Ahfeldt T, et al. (2010) Highly efficient reprogramming to pluripotency and directed differentiation of human cells with synthetic modified mRNA. Cell Stem Cell 7: 618-630. doi: 10.1016/j.stem.2010.08.012
|
| [17] |
Malik N, Rao MS (2013) A review of the methods for human iPSC derivation. Pluripotent Stem Cells: Methods and Protocols Totowa: Humana Press, 23-33. doi: 10.1007/978-1-62703-348-0_3
|
| [18] |
Hong H, Takahashi K, Ichisaka T, et al. (2009) Suppression of induced pluripotent stem cell generation by the p53–p21 pathway. Nature 460: 1132-1135. doi: 10.1038/nature08235
|
| [19] |
Zhao S, Jiang E, Chen S, et al. (2016) PiggyBac transposon vectors: the tools of the human gene encoding. Transl Lung Cancer Res 5: 120-125. doi: 10.21037/tcr.2016.04.02
|
| [20] |
Al Abbar A, Ngai SC, Nograles N, et al. (2020) Induced pluripotent stem cells: Reprogramming platforms and applications in cell replacement therapy. Biores Open Access 9: 121-136. doi: 10.1089/biores.2019.0046
|
| [21] |
Gouveia C, Huyser C, Egli D, et al. (2020) Lessons learned from somatic cell nuclear transfer. Int J Mol Sci 21: 2314. doi: 10.3390/ijms21072314
|
| [22] |
Tsuji Y, Kato Y, Tsunoda Y (2009) The developmental potential of mouse somatic cell nuclear-transferred oocytes treated with trichostatin A and 5-aza-2′-deoxycytidine. Zygote 17: 109-115. doi: 10.1017/S0967199408005133
|
| [23] |
Polstein LR, Perez-Pinera P, Kocak DD, et al. (2015) Genome-wide specificity of DNA binding, gene regulation, and chromatin remodeling by TALE- and CRISPR/Cas9-based transcriptional activators. Genome Res 25: 1158-1169. doi: 10.1101/gr.179044.114
|
| [24] |
Weltner J, Balboa D, Katayama S, et al. (2018) Human pluripotent reprogramming with CRISPR activators. Nat Commun 9: 2643. doi: 10.1038/s41467-018-05067-x
|
| [25] |
Liu P, Chen M, Liu Y, et al. (2017) CRISPR-based chromatin remodeling of the endogenous Oct4 or Sox2 locus enables reprogramming to pluripotency. Cell Stem Cell 22: 252-261.e4. doi: 10.1016/j.stem.2017.12.001
|
| [26] |
Ben JR, Shemer Y, Binah O (2018) Genome editing in induced pluripotent stem cells using CRISPR/Cas9. Stem Cell Rev Rep 14: 323-336. doi: 10.1007/s12015-018-9811-3
|
| [27] |
Brodehl A, Ebbinghaus H, Deutsch MA, et al. (2019) Human induced pluripotent stem-cell-derived cardiomyocytes as models for genetic cardiomyopathies. Int J Mol Sci 20: 4381. doi: 10.3390/ijms20184381
|
| [28] |
Kempf H, Zweigerdt R (2017) Scalable cardiac differentiation of pluripotent stem cells using specific growth factors and small molecules. Engineering and Application of Pluripotent Stem Cells Cham: Springer, 39-69. doi: 10.1007/10_2017_30
|
| [29] |
Wu S, Cheng CM, Lanz RB, et al. (2013) Atrial identity is determined by a COUP-TFII regulatory network. Dev Cell 25: 417-426. doi: 10.1016/j.devcel.2013.04.017
|
| [30] |
Prowse AB, Timmins NE, Yau TM, et al. (2014) Transforming the promise of pluripotent stem cell-derived cardiomyocytes to a therapy: challenges and solutions for clinical trials. Can J Cardiol 30: 1335-1349. doi: 10.1016/j.cjca.2014.08.005
|
| [31] |
Evans SM, Yelon D, Conlon FL, et al. (2010) Myocardial lineage development. Circ Res 107: 1428-1444. doi: 10.1161/CIRCRESAHA.110.227405
|
| [32] | Mahmood T, Nasser A, Hossein B (2015) Human cardiomyocyte generation from pluripotent stem cells: A state-of-art. Life Sciences 145: 98-113. |
| [33] |
Guo NN, Liu LP, Zheng YW, et al. (2020) Inducing human induced pluripotent stem cell differentiation through embryoid bodies: A practical and stable approach. J Stem Cells 12: 25-34. doi: 10.4252/wjsc.v12.i1.25
|
| [34] |
Brickman JM, Serup P (2017) Properties of embryoid bodies. WIREs Dev Biol 6: e259. doi: 10.1002/wdev.259
|
| [35] |
Yang L, Soonpaa MH, Adler ED, et al. (2008) Human cardiovascular progenitor cells develop from a KDR+ embryonic-stem-cell-derived population. Nature 453: 524-528. doi: 10.1038/nature06894
|
| [36] |
Kattman J, Witty AD, Gagliardi M, et al. (2011) Stage-specific optimization of activin/nodal and BMP signaling promotes cardiac differentiation of mouse and human pluripotent stem cell lines. Cell Stem Cell 8: 228-240. doi: 10.1016/j.stem.2010.12.008
|
| [37] |
Besser RR, Ishahak M, Mayo V, et al. (2018) Engineered microenvironments for maturation of stem cell derived cardiac myocytes. Theranostics 8: 124-140. doi: 10.7150/thno.19441
|
| [38] |
Tohyama S, Hattori F, Sano M, et al. (2012) Distinct metabolic flow enables large-scale purification of mouse and human pluripotent stem cell-derived cardiomyocytes. Cell Stem Cell 12: 127-137. doi: 10.1016/j.stem.2012.09.013
|
| [39] |
Mummery CL, Zhang J, Elliott DA, et al. (2012) Differentiation of human embryonic stem cells and induced pluripotent stem cells to cardiomyocytes: a methods overview. Circ Res 111: 344-358. doi: 10.1161/CIRCRESAHA.110.227512
|
| [40] |
Graichen R, Xu X, Braam SR, et al. (2008) Enhanced cardiomyogenesis of human embryonic stem cells by a small molecular inhibitor of p38 MAPK. Differentiation 76: 357-370. doi: 10.1111/j.1432-0436.2007.00236.x
|
| [41] |
Vlahos CJ, Matter WF, Hui KY, et al. (1994) A specific inhibitor of phosphatidylinositol3-kinase, 2-(4-morpholinyl)-8-phenyl-4H-1-benzopyran-4-one (LY294002). J Biol Chem 269: 5241-5248. doi: 10.1016/S0021-9258(17)37680-9
|
| [42] |
Ieda M, Fu JD, Delgado P, et al. (2010) Direct reprogramming of fibroblasts into functional cardiomyocytes by defined factors. Cell 142: 375-386. doi: 10.1016/j.cell.2010.07.002
|
| [43] |
Jayawardena TM, Egemnazarov B, Finch EA, et al. (2012) MicroRNA-mediated in vitro and in vivo direct reprogramming of cardiac fibroblasts to cardiomyocytes. Circ Res 110: 1465-1473. doi: 10.1161/CIRCRESAHA.112.269035
|
| [44] |
Burridge PW, Matsa E, Shukla P, et al. (2014) Chemically defined generation of human cardiomyocytes. Nat Methods 11: 855-860. doi: 10.1038/nmeth.2999
|
| [45] |
Srivastava D, DeWitt N (2016) In vivo cellular reprogramming: the next generation. Cell 166: 1386-1396. doi: 10.1016/j.cell.2016.08.055
|
| [46] |
Chen Y, Yang Z, Zhao ZA, et al. (2017) Direct reprogramming of fibroblasts into cardiomyocytes. Stem Cell Res Ther 8: 118. doi: 10.1186/s13287-017-0569-3
|
| [47] |
Wang J, Jiang X, Zhao L, et al. (2019) Lineage reprogramming of fibroblasts into induced cardiac progenitor cells by CRISPR/Cas9-based transcriptional activators. Acta Pharm Sin B 10: 313-326. doi: 10.1016/j.apsb.2019.09.003
|
| [48] |
Murphy SV, Atala A (2014) 3D bioprinting of tissues and organs. Nat Biotechnol 32: 773-785. doi: 10.1038/nbt.2958
|
| [49] |
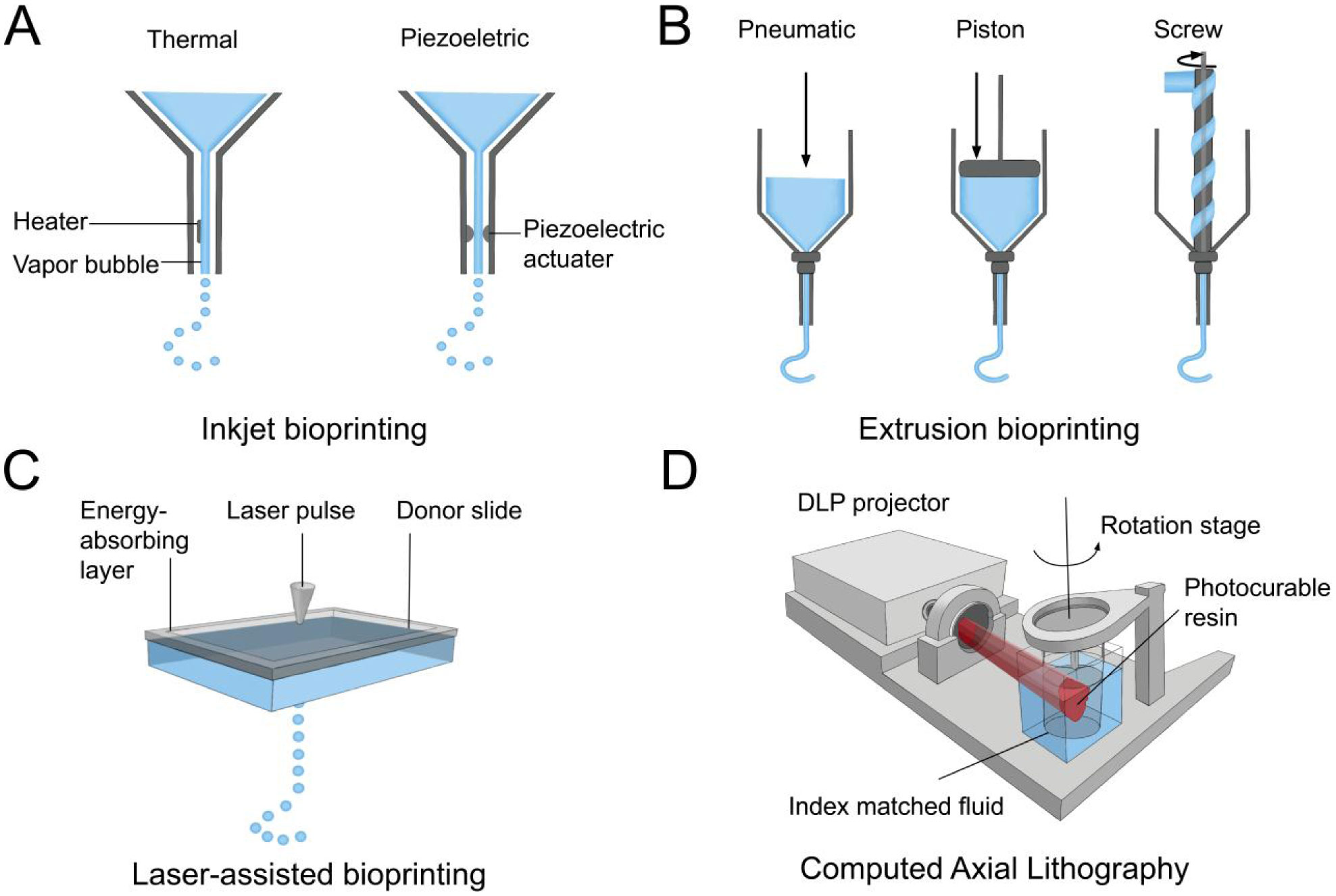
Cui X, Boland T, DLima DD, et al. (2012) Thermal inkjet printing in tissue engineering and regenerative medicine. Recent Pat Drug Deliv Formul 6: 149-155. doi: 10.2174/187221112800672949
|
| [50] |
Cui X, Dean D, Ruggeri ZM, et al. (2010) Cell damage evaluation of thermal inkjet printed Chinese hamster ovary cells. Biotechnol Bioeng 106: 963-969. doi: 10.1002/bit.22762
|
| [51] |
Fang Y, Frampton JP, Raghavan S, et al. (2012) Rapid generation of multiplexed cell cocultures using acoustic droplet ejection followed by aqueous two-phase exclusion patterning. Tissue Eng Part C-Me 18: 647-657. doi: 10.1089/ten.tec.2011.0709
|
| [52] |
Jones N (2012) Science in three dimensions: the print revolution. Nature 487: 22-23. doi: 10.1038/487022a
|
| [53] |
Mironov V, Visconti RP, Kasyanov V, et al. (2008) Organ printing: tissue spheroids as building blocks. Biomaterials 30: 2164-2174. doi: 10.1016/j.biomaterials.2008.12.084
|
| [54] |
Zhang YS, Pi Q, van Genderen AM Microfluidic bioprinting for engineering vascularized tissues and organoids (2017) . doi: 10.3791/55957
|
| [55] |
Bohandy J, Kim B, Adrian F (1986) Metal deposition from a supported metal film using an excimer laser. J Appl Phys 60: 1538-1539. doi: 10.1063/1.337287
|
| [56] |
Guillemot F, Souquet A, Catros S, et al. (2010) Laser-assisted cell printing: principle, physical parameters versus cell fate and perspectives in tissue engineering. Nanomedicine 5: 507-515. doi: 10.2217/nnm.10.14
|
| [57] |
Guillotin B, Souquet A, Catros S, et al. (2010) Laser assisted bioprinting of engineered tissue with high cell density and microscale organization. Biomaterials 31: 7250-7256. doi: 10.1016/j.biomaterials.2010.05.055
|
| [58] |
Kattamis NT, Purnick PE, Weiss R, et al. (2007) Thick film laser induced forward transfer for deposition of thermally and mechanically sensitive materials. Appl Phys Lett 91: 171120-171123. doi: 10.1063/1.2799877
|
| [59] |
Gauvin R, Chen YC, Lee JW, et al. (2012) Microfabrication of complex porous tissue engineering scaffolds using 3D projection stereolithography. Biomaterials 33: 3824-3834. doi: 10.1016/j.biomaterials.2012.01.048
|
| [60] |
Alonzo M, AnilKumar S, Roman B, et al. (2019) 3D Bioprinting of cardiac tissue and cardiac stem cell therapy. Transl Res 211: 64-83. doi: 10.1016/j.trsl.2019.04.004
|
| [61] |
Bishop ES, Mostafa S, Pakvasa M, et al. (2017) 3-D bioprinting technologies in tissue engineering and regenerative medicine: Current and future trends. Genes Dis 4: 185-195. doi: 10.1016/j.gendis.2017.10.002
|
| [62] |
Morris VB, Nimbalkar S, Younesi M, et al. (2017) Mechanical properties, cytocompatibility and manufacturability of chitosan: PEGDA hybrid-gel scaffolds by stereolithography. Ann Biomed Eng 45: 286-296. doi: 10.1007/s10439-016-1643-1
|
| [63] |
Lu Y, Mapili G, Suhali G, et al. (2006) A digital micro-mirror device-based system for the microfabrication of complex, spatially patterned tissue engineering scaffolds. J Biomed Mater Res A 77: 396-405. doi: 10.1002/jbm.a.30601
|
| [64] |
Zhang J, Hu Q, Wang S, et al. (2019) Digital light processing based three-dimensional printing for medical applications. Int J Bioprint 6: 242. doi: 10.18063/ijb.v6i1.242
|
| [65] |
Kelly BE, Bhattacharya I, Heidari H, et al. (2019) Volumetric additive manufacturing via tomographic reconstruction. Science 363: 1075-1079. doi: 10.1126/science.aau7114
|
| [66] |
Türker E, Demirçak N, Arslan YA (2018) Scaffold-free three-dimensional cell culturing using magnetic levitation. Biomater Sci 6: 1745-1753. doi: 10.1039/C8BM00122G
|
| [67] |
Matai I, Kaur G, Seyedsalehi A, et al. (2019) Progress in 3D bioprinting technology for tissue/organ regenerative engineering. Biomaterials 226: 119536. doi: 10.1016/j.biomaterials.2019.119536
|
| [68] |
Tseng H, Gage JA, Haisler WL, et al. (2016) A high-throughput in vitro ring assay for vasoactivity using magnetic 3D bioprinting. Sci Rep 6: 30640. doi: 10.1038/srep30640
|
| [69] |
Bowser DA, Moore MJ (2019) Biofabrication of neural microphysiological systems using magnetic spheroid bioprinting. Biofabrication 12: 015002. doi: 10.1088/1758-5090/ab41b4
|
| [70] |
Wang X (2019) Advanced polymers for three-dimensional (3D) organ bioprinting. Micromachines 10: 814. doi: 10.3390/mi10120814
|
| [71] |
Axpe E, Oyen ML (2016) Applications of alginate-based bioinks in 3D bioprinting. Int J Mol Sci 17: E1976. doi: 10.3390/ijms17121976
|
| [72] |
Bajpai SK, Sharma S (2004) Investigation of swelling/degradation behaviour of alginate beads crosslinked with Ca2+ and Ba2+ ions. React Funct Polym 59: 129-140. doi: 10.1016/j.reactfunctpolym.2004.01.002
|
| [73] |
Gao T, Gillispie GJ, Copus JS, et al. (2018) Optimization of gelatin–alginate composite bioink printability using rheological parameters: a systematic approach. Biofabrication 10: 034106. doi: 10.1088/1758-5090/aacdc7
|
| [74] |
Giuseppe MD, Law N, Webb BA, et al. (2018) Mechanical behaviour of alginate-gelatin hydrogels for 3D bioprinting. J Mech Behav Biomed Mater 79: 150-157. doi: 10.1016/j.jmbbm.2017.12.018
|
| [75] |
Markstedt K, Mantas A, Tournier I, et al. (2015) 3D bioprinting human chondrocytes with nanocellulose-alginate bioink for cartilage tissue engineering applications. Biomacromolecules 16: 1489-1496. doi: 10.1021/acs.biomac.5b00188
|
| [76] |
Frantz C, Stewart KM, Weaver VM (2010) The extracellular matrix at a glance. J Cell Sci 123: 4195-4200. doi: 10.1242/jcs.023820
|
| [77] |
Drzewiecki KE, Parmar AS, Gaudet ID, et al. (2014) Methacrylation induces rapid, temperature-dependent, reversible self-assembly of type-I collagen. Langmuir 30: 11204-11211. doi: 10.1021/la502418s
|
| [78] |
Lee A, Hudson AR, Shiwarski DJ, et al. (2019) 3D bioprinting of collagen to rebuild components of the human heart. Science 365: 482-487. doi: 10.1126/science.aav9051
|
| [79] |
Wang X, Yu X, Yan Y, et al. (2008) Liver tissue responses to gelatin and gelatin/chitosan gels. J Biomed Mater Res A 87: 62-68. doi: 10.1002/jbm.a.31712
|
| [80] |
Skardal A, Zhang J, McCoard L, et al. (2010) Photocrosslinkable hyaluronan-gelatin hydrogels for two-step bioprinting. Tissue Eng Part A 16: 2675-2685. doi: 10.1089/ten.tea.2009.0798
|
| [81] |
Zhu H, Yang H, Ma Y, et al. (2020) Spatiotemporally controlled photoresponsive hydrogels: design and predictive modeling from processing through application. Adv Funct Mater 30: 2000639. doi: 10.1002/adfm.202000639
|
| [82] |
Xiao S, Zhao T, Wang J, et al. (2019) Gelatin methacrylate (GelMA)-based hydrogels for cell transplantation: an effective strategy for tissue engineering. Stem Cell Rev Rep 15: 664-679. doi: 10.1007/s12015-019-09893-4
|
| [83] |
Hoffman AS (2002) Hydrogels for biomedical applications. Adv Drug Deliv Rev 54: 3-12. doi: 10.1016/S0169-409X(01)00239-3
|
| [84] |
Jungst T, Smolan W, Schacht K, et al. (2016) Strategies and molecular design criteria for 3D printable hydrogels. Chem Rev 116: 1496-1539. doi: 10.1021/acs.chemrev.5b00303
|
| [85] |
Astete CE, Sabliov CM (2006) Synthesis and characterization of PLGA nanoparticles. J Biomater Sci Polym Ed 17: 247-289. doi: 10.1163/156856206775997322
|
| [86] |
Samadi N, Abbadessa A, Di Stefano A, et al. (2013) The effect of lauryl capping group on protein release and degradation of poly (D, L-lactic-co-glycolic acid) particles. J Control Release 172: 436-443. doi: 10.1016/j.jconrel.2013.05.034
|
| [87] |
Mazzola M, Pasquale E (2020) Toward cardiac regeneration: Combination of pluripotent stem cell-based therapies and bioengineering strategies. Front Bioeng Biotechnol 8: 455. doi: 10.3389/fbioe.2020.00455
|
| [88] |
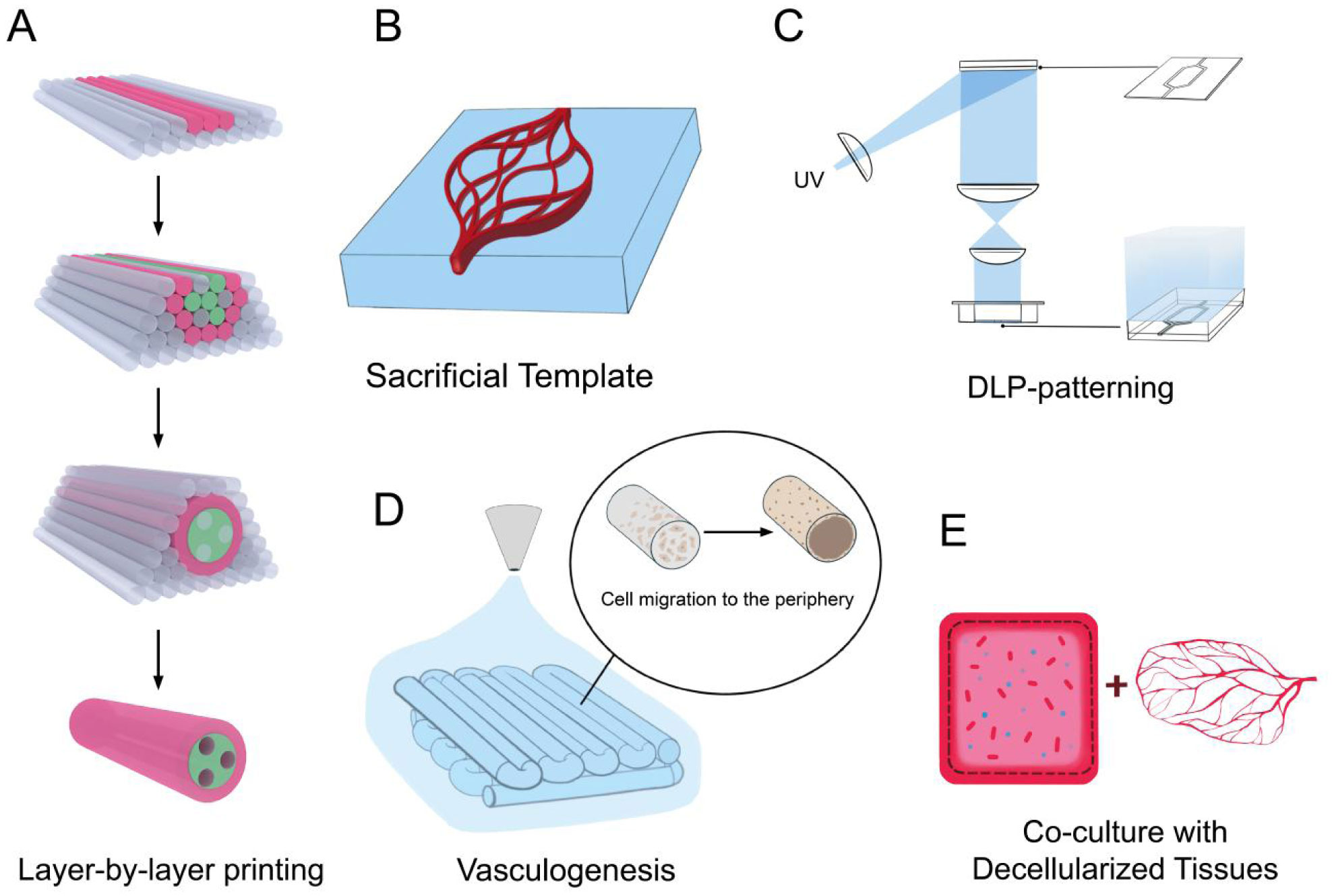
Homma J, Shimizu S, Sekine H, et al. (2020) A novel method to align cells in a cardiac tissue-like construct fabricated by cell sheet-based tissue engineering. J Tissue Eng Regen Med 14: 944-954. doi: 10.1002/term.3074
|
| [89] |
Wang Z, Lee SJ, Cheng HJ, et al. (2018) 3D bioprinted functional and contractile cardiac tissue constructs. Acta Biomater 70: 48-56. doi: 10.1016/j.actbio.2018.02.007
|
| [90] |
Redd MA, Zeinstra N, Qin W, et al. (2019) Patterned human microvascular grafts enable rapid vascularization and increase perfusion in infarcted rat hearts. Nat Commun 10: 584. doi: 10.1038/s41467-019-08388-7
|
| [91] |
Shimizu A, Goh WH, Itai S, et al. (2020) ECM-based microchannel for culturing in vitro vascular tissues with simultaneous perfusion and stretch. Lab Chip 20: 1917-1927. doi: 10.1039/D0LC00254B
|
| [92] |
Ma X, Qu X, Zhu W, et al. (2016) Deterministically patterned biomimetic human iPSC-derived hepatic model via rapid 3D bioprinting. Proc Natl Acad Sci 113: 2206-2211. doi: 10.1073/pnas.1524510113
|
| [93] |
Taylor DA, Sampaio LC, Ferdous Z, et al. (2018) Decellularized matrices in regenerative medicine. Acta Biomater 74: 74-89. doi: 10.1016/j.actbio.2018.04.044
|
| [94] |
Ott HC, Matthiesen TS, Goh SK, et al. (2008) Perfusion-decellularized matrix: using nature's platform to engineer a bioartificial heart. Nat Med 14: 213-219. doi: 10.1038/nm1684
|
| [95] |
Jovic TH, Kungwengwe G, Mills AC, et al. (2019) Plant-derived biomaterials: A review of 3D bioprinting and biomedical applications. Front Mech Eng 5: 19. doi: 10.3389/fmech.2019.00019
|
| [96] |
Marga F, Jakab K, Khatiwala C, et al. (2012) Toward engineering functional organ modules by additive manufacturing. Biofabrication 4: 022001. doi: 10.1088/1758-5082/4/2/022001
|
| [97] |
Hockaday LA, Kang KH, Colangelo NW, et al. (2012) Rapid 3D printing of anatomically accurate and mechanically heterogeneous aortic valve hydrogel scaffolds. Biofabrication 4: 035005. doi: 10.1088/1758-5082/4/3/035005
|
| [98] |
Madden LR, Mortisen DJ, Sussman EM, et al. (2010) Proangiogenic scaffolds as functional templates for cardiac tissue engineering. Proc Natl Acad Sci 107: 15211-15216. doi: 10.1073/pnas.1006442107
|
| [99] |
Zhang YS, Arneri A, Bersini S, et al. (2016) Bioprinting 3D microfibrous scaffolds for engineering endothelialized myocardium and heart-on-a-chip. Biomaterials 110: 45-59. doi: 10.1016/j.biomaterials.2016.09.003
|
| [100] |
Hann SY, Cui H, Esworthy T, et al. (2019) Recent advances in 3D printing: vascular network for tissue and organ regeneration. Transl Res 211: 46-63. doi: 10.1016/j.trsl.2019.04.002
|
| [101] |
Duan B, Hockaday LA, Kang KH, et al. (2013) 3D bioprinting of heterogeneous aortic valve conduits with alginate/gelatin hydrogels. J Biomed Mater Res A 101: 1255-1264. doi: 10.1002/jbm.a.34420
|
| [102] |
Grigoryan B, Paulsen SJ, Corbett DC, et al. (2019) Multivascular networks and functional intravascular topologies within biocompatible hydrogels. Science 364: 458-464. doi: 10.1126/science.aav9750
|
| [103] |
Esther C, Claudia K, Petra J (2011) Vascularization is the key challenge in tissue engineering. Adv Drug Deliv Rev 63: 300-311. doi: 10.1016/j.addr.2011.03.004
|
| [104] |
Kobayashi J, Akiyama Y, Yamato M, et al. (2018) Design of temperature-responsive cell culture surfaces for cell sheet-based regenerative therapy and 3D tissue fabrication. Adv Exp Med Biol 1078: 371-393. doi: 10.1007/978-981-13-0950-2_19
|
| [105] |
Inui A, Sekine H, Sano K, et al. (2019) Generation of a large-scale vascular bed for the in vitro creation of three-dimensional cardiac tissue. Regen Ther 11: 316-323. doi: 10.1016/j.reth.2019.10.001
|
| [106] |
Masuda S, Shimizu T (2015) Three-dimensional cardiac tissue fabrication based on cell sheet technology. Adv Drug Deliv Rev 96: 103-109. doi: 10.1016/j.addr.2015.05.002
|
| [107] |
Daley MC, Fenn SL, Black LD (2018) Applications of cardiac extracellular matrix in tissue engineering and regenerative medicine. Cardiac Extracellular Matrix Cham: Springer, 59-83. doi: 10.1007/978-3-319-97421-7_4
|
| [108] |
Seignez C, Phillipson M (2017) The multitasking neutrophils and their involvement in angiogenesis. Curr Opin Hematol 24: 3-8. doi: 10.1097/MOH.0000000000000300
|
| [109] |
Miller JS (2014) The billion cells construct: will three-dimensional printing get us there? PLoS Biol 12: e1001882. doi: 10.1371/journal.pbio.1001882
|
Figures(3)

Yihan Zhang. Manufacture of complex heart tissues: technological advancements and future directions[J]. AIMS Bioengineering, 2021, 8(1): 73-92. doi: 10.3934/bioeng.2021008







 DownLoad:
DownLoad: