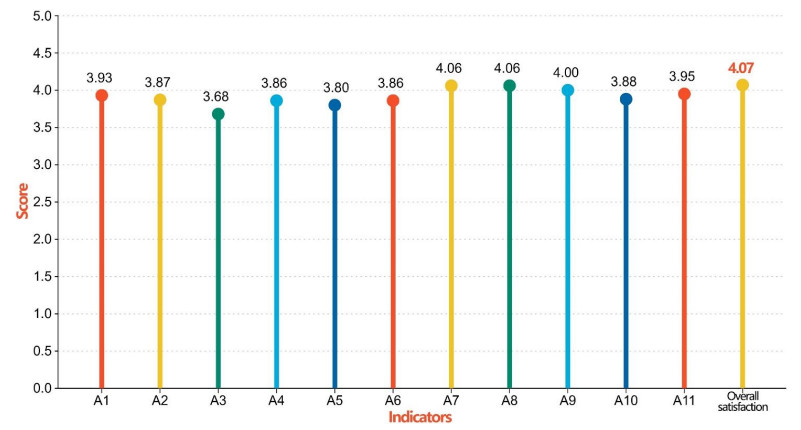
The cultural landscape of traditional villages is a valuable cultural heritage. Using the example of the Huangdu Dong Village, this study collected data on the perceptions of 209 tourists regarding the cultural landscape of traditional villages using a questionnaire survey. The perceptions and satisfaction rates of tourists were analyzed, and key factors influencing their satisfaction with the cultural landscape were identified. The results show that tourists generally hold a positive perception of traditional village cultural landscapes, with the highest levels of perceptions belonging to folk activity cultural landscapes and clothing cultural landscapes. Additionally, this study identified four key factors that have a significant positive impact on tourist satisfaction with the cultural landscape, namely architectural, water, vegetation, and service facility cultural landscapes. The research also found that tourists residing in rural areas reported significantly higher satisfaction with the cultural landscape than those living in urban areas; moreover, tourists with an income above 10,000 yuan exhibited significantly higher satisfaction compared to other income groups. Finally, based on its findings, this study provides recommendations for optimizing the protection, planning, and design of traditional village cultural landscapes to enhance overall tourist satisfaction.
Citation: Huaheng Shen, Xueqin Tan, Xinmei Liu, Xiting Yu, Yu Luo. Perceptions of cultural landscapes: Exploring tourist satisfaction in traditional villages[J]. AIMS Geosciences, 2025, 11(1): 7-26. doi: 10.3934/geosci.2025002
The cultural landscape of traditional villages is a valuable cultural heritage. Using the example of the Huangdu Dong Village, this study collected data on the perceptions of 209 tourists regarding the cultural landscape of traditional villages using a questionnaire survey. The perceptions and satisfaction rates of tourists were analyzed, and key factors influencing their satisfaction with the cultural landscape were identified. The results show that tourists generally hold a positive perception of traditional village cultural landscapes, with the highest levels of perceptions belonging to folk activity cultural landscapes and clothing cultural landscapes. Additionally, this study identified four key factors that have a significant positive impact on tourist satisfaction with the cultural landscape, namely architectural, water, vegetation, and service facility cultural landscapes. The research also found that tourists residing in rural areas reported significantly higher satisfaction with the cultural landscape than those living in urban areas; moreover, tourists with an income above 10,000 yuan exhibited significantly higher satisfaction compared to other income groups. Finally, based on its findings, this study provides recommendations for optimizing the protection, planning, and design of traditional village cultural landscapes to enhance overall tourist satisfaction.

| [1] | Gao J, Wu B, Zhao Z (2020) Construction of Sustainable Path Model for Traditional Village Tourism Revitalization Based on the Perspective of Cultural Geography. Regional Res Dev 39: 73–78. |
| [2] | Tang C, Wan Z, Liu M, et al. (2021) Multi-agent-based Traditional Village Cultural Heritage Protection, Inheritance, Perception and Improvement Model. Arid Area Resour Environ 35: 196–202. |
| [3] |
Wu C, Chen M, Zhou L, et al. (2020) Identifying the Spatiotemporal Patterns of Traditional Villages in China: A Multiscale Perspective. Land 9: 449. https://doi.org/10.3390/land9110449 doi: 10.3390/land9110449

|
| [4] |
Chen Z, Yang H, Lin Y, et al. (2024) Exploring the association between the built environment and positive sentiments of tourists in traditional villages in Fuzhou, China. Ecol Inf 80: 102465. https://doi.org/10.1016/j.ecoinf.2024.102465 doi: 10.1016/j.ecoinf.2024.102465
|
| [5] |
Shen J, Chou RJ (2022) Rural revitalization of Xiamei: The development experiences of integrating tea tourism with ancient village preservation. J Rural Stud 90: 42–52. https://doi.org/10.1016/j.jrurstud.2022.01.006 doi: 10.1016/j.jrurstud.2022.01.006
|
| [6] |
Wang F, Zhao X, Qiu Y, et al. (2023) Adaptability of traditional villages as tourist destinations in Yellow River Basin, China. Indoor Built Environ 32: 574–589. https://doi.org/10.1177/1420326X221126056 doi: 10.1177/1420326X221126056
|
| [7] |
Ćurčić N, Mirković Svitlica A, Brankov J, et al. (2021) The Role of Rural Tourism in Strengthening the Sustainability of Rural Areas: The Case of Zlakusa Village. Sustainability 13: 6747. https://doi.org/10.3390/su13126747 doi: 10.3390/su13126747
|
| [8] |
Kataya A (2021) The Impact of Rural Tourism on the Development of Regional Communities. J East Europe Res Bus Econ 2021: 1–10. https://doi.org/10.5171/2021.652463 doi: 10.5171/2021.652463
|
| [9] |
Nematpour M, Khodadadi M (2021) Farm tourism as a driving force for socioeconomic development: a benefits viewpoint from Iran. Curr Issues Tour 24: 247–263. https://doi.org/10.1080/13683500.2020.1711712 doi: 10.1080/13683500.2020.1711712
|
| [10] |
Vu Dinh H, Ngo Anh T, Nguyen Thi Ngoc A, et al. (2023) Strategies for Sustainable Rural Tourism Innovation: Evidence from Hanoi, Vietnam. J Environ Manag Tour 14: 1984. https://doi.org/10.14505/jemt.v14.4(68).10 doi: 10.14505/jemt.v14.4(68).10
|
| [11] | Ruhanen L, Whitford M (2021) Cultural heritage and Indigenous tourism, Indigenous heritage, Routledge, 1–13. |
| [12] |
Li X, Wang C (2023) Understanding the relationship between tourists' perceptions of the authenticity of traditional village cultural landscapes and behavioural intentions, mediated by memorable tourism experiences and place attachment. Asia Pac J Tour Res 28: 254–273. https://doi.org/10.1080/10941665.2023.2217959 doi: 10.1080/10941665.2023.2217959
|
| [13] |
Tieskens KF, Schulp CJE, Levers C, et al. (2017) Characterizing European cultural landscapes: Accounting for structure, management intensity and value of agricultural and forest landscapes. Land Use Policy 62: 29–39. https://doi.org/10.1016/j.landusepol.2016.12.001 doi: 10.1016/j.landusepol.2016.12.001
|
| [14] |
Job H, Paesler F (2013) Links between nature-based tourism, protected areas, poverty alleviation and crises—The example of Wasini Island (Kenya). J Outdoor Rec Tour 1–2: 18–28. https://doi.org/10.1016/j.jort.2013.04.004 doi: 10.1016/j.jort.2013.04.004
|
| [15] |
Gocer O, Boyacioglu D, Karahan EE, et al. (2024) Cultural tourism and rural community resilience: A framework and its application. J Rural Stud 107: 103238. https://doi.org/10.1016/j.jrurstud.2024.103238 doi: 10.1016/j.jrurstud.2024.103238
|
| [16] |
Kang N, Liu C (2024) Assessment of visual quality and social perception of cultural landscapes: application to Anyi traditional villages, China. Herit Sci 12: 235. https://doi.org/10.1186/s40494-024-01326-x doi: 10.1186/s40494-024-01326-x
|
| [17] |
Shen H, Aziz NF, Liu J, et al. (2024) From text to insights: leveraging NLP to assess how landscape features shape tourist perceptions and emotions toward traditional villages. Environ Res Commun 6: 115006. https://doi.org/10.1088/2515-7620/ad8ca3 doi: 10.1088/2515-7620/ad8ca3
|
| [18] |
Shen H, Aziz NF, Omar SI, et al. (2024) Tourism's impact on visual landscape: Residents' perceptions from a traditional Chinese village. Pol J Environ Stud 33: 4707–4719. https://doi.org/10.15244/pjoes/176229 doi: 10.15244/pjoes/176229
|
| [19] |
Amalia R, Wibisono N, Elliott-White M (2023) Increasing Tourist Revisit Intention in Garut Tourist Attractions: The Role of Destination Image and Tourist Satisfaction. JMI 3. https://doi.org/10.35313/jmi.v3i2.74 doi: 10.35313/jmi.v3i2.74
|
| [20] |
Luvsandavaajav O, Narantuya G, Dalaibaatar E, et al. (2022) A longitudinal study of destination image, tourist satisfaction, and revisit intention. J Tour Serv 13: 128–149. https://doi.org/10.29036/jots.v13i24.341 doi: 10.29036/jots.v13i24.341
|
| [21] |
Huete Alcocer N, López Ruiz VR (2020) The role of destination image in tourist satisfaction: the case of a heritage site. Econ Res-Ekon Istraž 33: 2444–2461. https://doi.org/10.1080/1331677X.2019.1654399 doi: 10.1080/1331677X.2019.1654399
|
| [22] |
Shen H, Aziz NF, Huang M, et al. (2023) Impact of the Perceived Quality of Traditional Villages' Cultural Landscapes on Tourists' Loyalty. J Environ Manag Tour 14: 2853–2870. https://doi.org/10.14505/jemt.v14.7(71).04 doi: 10.14505/jemt.v14.7(71).04
|
| [23] |
Caravello E (2023) Preserving cultural landscapes in the face of globalization. The musealization of Sicilian heritage. AIMS Geosci 9: 697–712. https://doi.org/10.3934/geosci.2023037 doi: 10.3934/geosci.2023037
|
| [24] |
Xu W, Zhao J, Ye L (2018) Culture is new nature: comparing the restorative capacity of cultural and natural landscapes. Int J Environ Stud 75: 847–865. https://doi.org/10.1080/00207233.2018.1426311 doi: 10.1080/00207233.2018.1426311
|
| [25] |
Tieskens KF, Van Zanten BT, Schulp CJE, et al. (2018) Aesthetic appreciation of the cultural landscape through social media: An analysis of revealed preference in the Dutch river landscape. Landscape Urban Plan 177: 128–137. https://doi.org/10.1016/j.landurbplan.2018.05.002 doi: 10.1016/j.landurbplan.2018.05.002
|
| [26] |
Yang W, Fan B, Tan J, et al. (2022) The spatial perception and spatial feature of rural cultural landscape in the context of rural tourism. Sustainability 14: 4370. https://doi.org/10.3390/su14074370 doi: 10.3390/su14074370
|
| [27] |
Zhou J, Wu S, Wu X, et al. (2023) Cultural landscape perception of the Chinese traditional settlement: Based on tourists' online comments. PLoS ONE 18: e0283335. https://doi.org/10.1371/journal.pone.0283335 doi: 10.1371/journal.pone.0283335
|
| [28] |
Lu J, Ma C, Ma C (2020) Landscape evaluation of traditional villages based on AHP-GIS analysis. Jiangsu Agric Sci 48: 298–306. https://doi.org/10.15889/j.issn.1002-1302.2020.10.055 doi: 10.15889/j.issn.1002-1302.2020.10.055
|
| [29] | Sauer C (2008) The morphology of landscape, The cultural geography reader, Routledge, 108–116. |
| [30] | World Heritage Committee (2002) Operational Guidelines for the Implementation of the World Heritage Convention. https://coilink.org/20.500.12592/tb2rc6m. |
| [31] |
Xu Q, Wang J (2021) Recognition of values of traditional villages in southwest China for sustainable development: A case study of Liufang Village. Sustainability 13: 7569. https://doi.org/10.3390/su13147569 doi: 10.3390/su13147569
|
| [32] |
Kang N, Xie G, Liu C (2023) Assessment of Society's Perceptions on Cultural Ecosystem Services in a Cultural Landscape in Nanchang, China. Sustainability 15: 10308. https://doi.org/10.3390/su151310308 doi: 10.3390/su151310308
|
| [33] | Shuib KB, Hashim H (2011) Cultural landscape values of a rural landscape: Perception of outsiders and tourists, International Federation of Landscape Architects Asia-Pacific Region (IFLA APR) Congress: Hospitality. Available from: https://www.academia.edu/download/70014267/OR026.pdf. |
| [34] |
Santoro A, Venturi M, Agnoletti M (2021) Landscape Perception and Public Participation for the Conservation and Valorization of Cultural Landscapes: The Case of the Cinque Terre and Porto Venere UNESCO Site. Land 10: 93. https://doi.org/10.3390/land10020093 doi: 10.3390/land10020093
|
| [35] |
Cheng G, Li Z, Xia S, et al. (2023) Research on the Spatial Sequence of Building Facades in Huizhou Regional Traditional Villages. Buildings 13: 174. https://doi.org/10.3390/buildings13010174 doi: 10.3390/buildings13010174
|
| [36] |
Dragan A, Crețan R, Terian MI (2024) Landscapes of Watermills: A Rural Cultural Heritage Perspective in an East-Central European Context. Heritage 7: 4790–4813. https://doi.org/10.3390/heritage7090227 doi: 10.3390/heritage7090227
|
| [37] | Mbaiwa JE, Siphambe GB (2023) Rural Heritage and Tourism in Africa, Cultural Heritage and Tourism in Africa, Routledge, 116–136. |
| [38] |
Moscatelli M (2024) Heritage as a Driver of Sustainable Tourism Development: The Case Study of the Darb Zubaydah Hajj Pilgrimage Route. Sustainability 16: 7055. https://doi.org/10.3390/su16167055 doi: 10.3390/su16167055
|
| [39] |
Parta IBMW, Maharani IAK (2023) CULTURAL TOURISM IN INDONESIA: SYSTEMATIC LITERATURE REVIEW. Vidyottama Sanatana Int J Hindu Sci Religious Stud 7: 189–204. https://doi.org/10.25078/vidyottama.v7i2.2498 doi: 10.25078/vidyottama.v7i2.2498
|
| [40] |
Yang L, Hu Y, Wu X, et al. (2023) Cultural landscape gene production process and mechanism of Dong traditional village: A case study of Huangdu village. J Nat Resour 38: 1164–1177. https://doi.org/10.31497/zrzyxb.20230504 doi: 10.31497/zrzyxb.20230504
|
| [41] |
Tian M, Zheng W, Wang N (2021) Research on the Interactive Development of Dong Village Cultural Heritage Protection and Night Tourism: a Case Study of Huangdu Dong Village in Hunan. E3S Web Conf 251: 02005. https://doi.org/10.1051/e3sconf/202125102005 doi: 10.1051/e3sconf/202125102005
|
| [42] |
Zhang Q, Kim E, Yang C, et al. (2023) Rural revitalization: sustainable strategy for the development of cultural landscape of traditional villages through optimized IPA approach. J Cult Herit Manag S 13: 66–86. https://doi.org/10.1108/JCHMSD-09-2020-0130 doi: 10.1108/JCHMSD-09-2020-0130
|
| [43] | Brown JD (2011) Likert items and scales of measurement. Statistics 15: 10–14. |
| [44] |
Zhang L, Miao Y, Linna A (2023) An Experimental Eye-Movement Study on the Spatial Attraction Characteristics and Perception of Different Landscape Types in Ethnic Villages. J Urban Plann Dev 149: 04023039. https://doi.org/10.1061/JUPDDM.UPENG-4348 doi: 10.1061/JUPDDM.UPENG-4348
|
| [45] |
Yang Z, Chen J, Duan J, et al. (2021) Evaluation of vernacular landscape attraction for traditional village tourism: A case study of Yongfeng Village in Lanzhou. E3S Web Conf 292: 03058. https://doi.org/DOI:10.1051/e3sconf/202129203058 doi: 10.1051/e3sconf/202129203058
|
| [46] |
Chen H, Tao W (2017) The Revival and Restructuring of a Traditional Folk Festival: Cultural Landscape and Memory in Guangzhou, South China. Sustainability 9: 1767. https://doi.org/10.3390/su9101767 doi: 10.3390/su9101767
|
| [47] | He Y, Thawascha Dechsubha Z (2023) Visitors' perceived symbol, experience and behavioral intention in folk festival: take Guangfu Temple Fair an example. J Surv Fish Sci 10: 1442–1452. |
| [48] |
Zhuang Q, Wan M, Zheng G (2022) Presentation and Elaboration of the Folk Intangible Cultural Heritage from the Perspective of the Landscape. Buildings 12: 1388. https://doi.org/10.3390/buildings12091388 doi: 10.3390/buildings12091388
|
| [49] | Bohórquez E, Pérez M, Benavides A, et al. (2021) Folklore and Tourism: Folk Dance Groups as a Strategy to Promote the Development of Cultural Tourism, Advances in Tourism, Technology and Systems, ICOTTS 2020. Smart Innovation, Systems and Technologies, Springer, Singapore. 2: 106–113. https://doi.org/10.1007/978-981-33-4260-6_10 |
| [50] | Liu Z, Hanisa H, Yang Y (2024) An Aesthetic Morphology Approach of Visual Identity on Dong's Ethnic Costume. Asian J Res Educ Soc Sci 6: 31–43. |
| [51] |
Ni J (2020) Tourism Value of Traditional Costume Culture in Ethnic Tourism Villages. Creativity Innovation 4: 38–47. https://doi.org/10.47297/wspciWSP2516-252705.20200406 doi: 10.47297/wspciWSP2516-252705.20200406
|
| [52] |
Guo Z, Sun L (2016) The planning, development and management of tourism: The case of Dangjia, an ancient village in China. Tourism Manage 56: 52–62. https://doi.org/10.1016/j.tourman.2016.03.017 doi: 10.1016/j.tourman.2016.03.017
|
| [53] |
Xu L, Sang K, Li G, et al. (2023) Heritage evaluation and analysis based on entropy weight method: the study of Wengji ancient village in China. J Hous Built Environ 38: 1843–1868. https://doi.org/10.1007/s10901-023-10019-z doi: 10.1007/s10901-023-10019-z
|
| [54] |
Joshi NC, Rawat GS (2020) Why only blue in the traditional architecture of western Himalaya, India? Color Res Appl 45: 95–104. https://doi.org/10.1002/col.22440 doi: 10.1002/col.22440
|
| [55] |
Asur F (2019) An evaluation of visual landscape quality of coastal settlements: A case study of coastal areas in the Van Lake Basin (Turkey). Appl Ecol Env Res 17: 1849–1864. http://dx.doi.org/10.15666/aeer/1702_18491864 doi: 10.15666/aeer/1702_18491864
|
| [56] |
Yamashita S (2002) Perception and evaluation of water in landscape: use of Photo-Projective Method to compare child and adult residents' perceptions of a Japanese river environment. Landscape Urban Plan 62: 3–17. https://doi.org/10.1016/S0169-2046(02)00093-2 doi: 10.1016/S0169-2046(02)00093-2
|
| [57] |
Cai K, Huang W, Lin G (2022) Bridging landscape preference and landscape design: A study on the preference and optimal combination of landscape elements based on conjoint analysis. Urban For Urban Gree 73: 127615. https://doi.org/10.1016/j.ufug.2022.127615 doi: 10.1016/j.ufug.2022.127615
|
| [58] |
Shen H, Aziz NF, Huang M, et al. (2024) Tourist perceptions of landscape in Chinese traditional villages: analysis based on online data. J Tour Cult Change 22: 232–251. https://doi.org/10.1080/14766825.2023.2240297 doi: 10.1080/14766825.2023.2240297
|
| [59] | Perera NF, Perera ERK, Perera ANK (2009) Traditional village-based hydraulic culture. Economic review June/July. |
| [60] |
Yodsurang P, Hiromi M, Yasufumi U (2015) A traditional community in the Chao Phraya River Basin Ⅱ: Influence of water circulation on the traditional living culture according to the settlement pattern. Asian Cult Hist 8: 112–125. https://doi.org/10.5539/ach.v8n1p57 doi: 10.5539/ach.v8n1p57
|
| [61] |
Jeon JY, Jo HI (2020) Effects of audio-visual interactions on soundscape and landscape perception and their influence on satisfaction with the urban environment. Build Environ 169: 106544. https://doi.org/10.1016/j.buildenv.2019.106544 doi: 10.1016/j.buildenv.2019.106544
|
| [62] |
Zhao J, Xu W, Ye L (2018) Effects of auditory-visual combinations on perceived restorative potential of urban green space. Appl Acoust 141: 169–177. https://doi.org/10.1016/j.apacoust.2018.07.001 doi: 10.1016/j.apacoust.2018.07.001
|
| [63] | Tang S, Wang C, Zhang C, et al. (2018) Plant landscape characteristics and the impact of village expansion on it in three traditional villages of Guangzhou, southern China. J Beijing For Univ 40: 90–102. |
| [64] | Sujarwo W (2019) Sasak Traditional Villages: A tourism potential and conservation effort for culture and plants. J Masyarakat Budaya 21: 203–220. |
| [65] |
Zhuang X, Du B (2024) Cultural Implication and the Application of Native Plants under the Perspective of Cultural Ecology in Rural Revitalization. Chin Overseas Archit, 109–114. https://doi.org/DOI:10.19940/j.cnki.1008-0422.2024.03.018 doi: 10.19940/j.cnki.1008-0422.2024.03.018
|
| [66] |
Polat AT, Akay A (2015) Relationships between the visual preferences of urban recreation area users and various landscape design elements. Urban For Urban Gree 14: 573–582. https://doi.org/10.1016/j.ufug.2015.05.009 doi: 10.1016/j.ufug.2015.05.009
|
| [67] |
Gobster PH, Westphal LM (2004) The human dimensions of urban greenways: planning for recreation and related experiences. Landscape Urban Plan 68: 147–165. https://doi.org/doi:10.1016/S0169-2046(03)00162-2 doi: 10.1016/S0169-2046(03)00162-2
|
| [68] |
Jiang S, Ma H, Yang L, et al. (2023) The Influence of Perceived Physical and Aesthetic Quality of Rural Settlements on Tourists' Preferences—A Case Study of Zhaoxing Dong Village. Land 12: 1542. https://doi.org/10.3390/land12081542 doi: 10.3390/land12081542
|
| [69] |
Oviedo-García MÁ, Vega-Vázquez M, Castellanos-Verdugo M, et al. (2019) Tourism in protected areas and the impact of servicescape on tourist satisfaction, key in sustainability. J Destin Mark Manag 12: 74–83. https://doi.org/10.1016/j.jdmm.2019.02.005 doi: 10.1016/j.jdmm.2019.02.005
|
| [70] |
Wang C, Hao Y (2023) Empirical analysis of tourist satisfaction of leisure farms: evidence from Qing Jing Farms, Taiwan. Humanit Soc Sci Commun 10: 1–9. https://doi.org/10.1057/s41599-023-01901-w doi: 10.1057/s41599-023-01901-w
|
| [71] |
Wang S, Wang J, Shen W, et al. (2023) The evaluation of tourism service facilities in Chinese traditional villages based on the living protection concept: theoretical framework and empirical case study. J Asian Archit Build 22: 14–31. https://doi.org/10.1080/13467581.2021.2007109 doi: 10.1080/13467581.2021.2007109
|
| [72] |
Liang N, Maulan S, Yusof MJM, et al. (2023) The Relationship of Visual Preference for Chinese Traditional Village Landscape with Perceived Values and Functions. AJACT 5. https://doi.org/10.55057/ajact.2023.5.3.2 doi: 10.55057/ajact.2023.5.3.2
|
| [73] |
Carrascal Incera A, Fernández MF (2015) Tourism and income distribution: Evidence from a developed regional economy. Tourism Manage 48: 11–20. https://doi.org/10.1016/j.tourman.2014.10.016 doi: 10.1016/j.tourman.2014.10.016
|
| [74] |
Waqas-Awan A, Rosselló-Nadal J, Santana-Gallego M (2021) New Insights into the Role of Personal Income on International Tourism. J Travel Res 60: 799–809. https://doi.org/10.1177/0047287520907702 doi: 10.1177/0047287520907702
|
| [75] |
Taheri B, Gannon MJ, Kesgin M (2020) Visitors' perceived trust in sincere, authentic, and memorable heritage experiences. Serv Ind J 40: 705–725. https://doi.org/10.1080/02642069.2019.1642877 doi: 10.1080/02642069.2019.1642877
|
| [76] | Gu H, Ryan C (2014) Hongcun, china—residents' perceptions of the impacts of tourism on a rural community: a mixed methods approach, Contemporary Perspectives on China Tourism, Routledge, 177–204. |


Figures(4) / Tables(8)

Huaheng Shen, Xueqin Tan, Xinmei Liu, Xiting Yu, Yu Luo. Perceptions of cultural landscapes: Exploring tourist satisfaction in traditional villages[J]. AIMS Geosciences, 2025, 11(1): 7-26. doi: 10.3934/geosci.2025002







 DownLoad:
DownLoad: