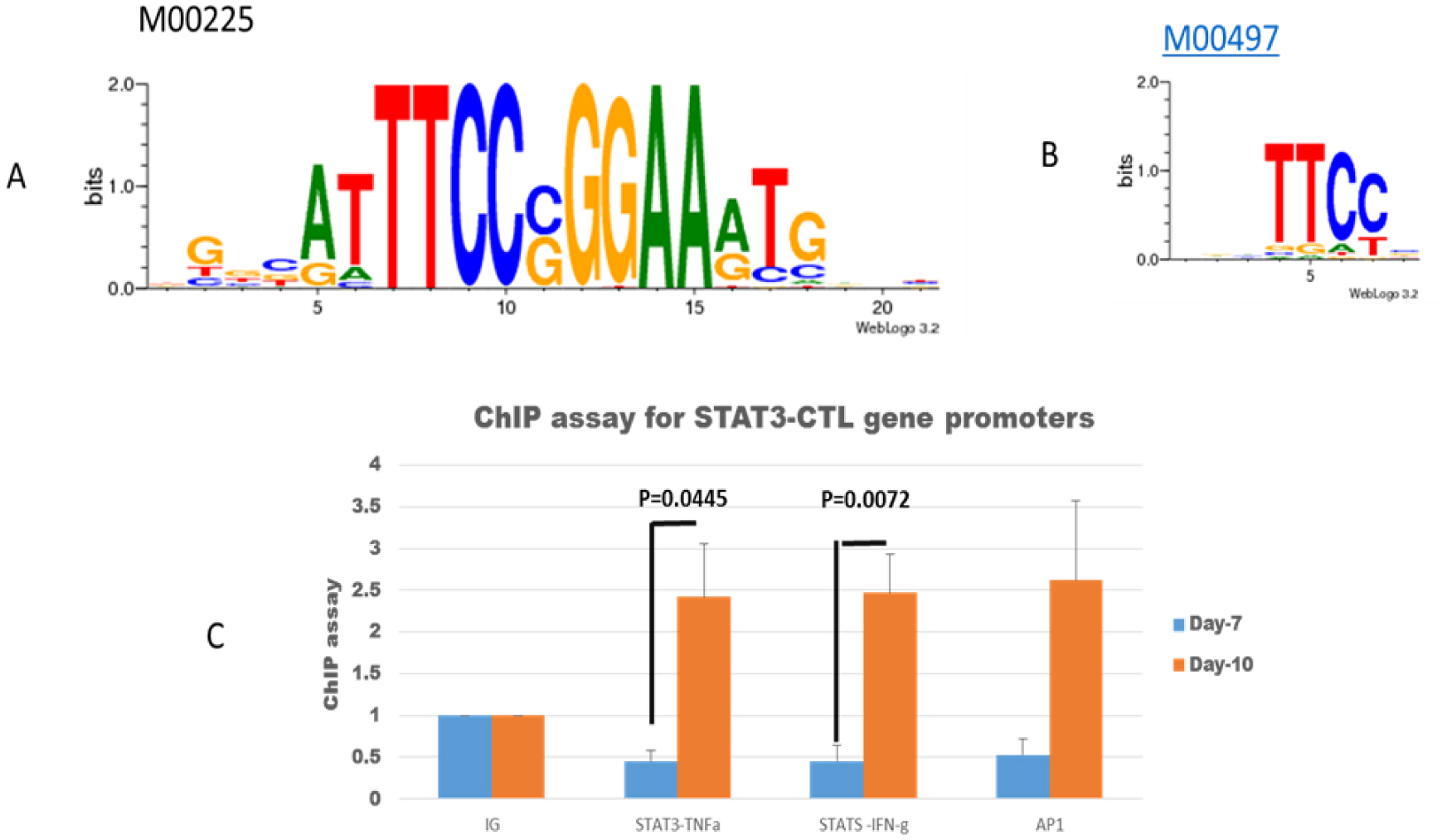
Tumor-infiltrating lymphocytes (TILs) can be used in two clinical areas: Adoptive cell therapy (ACT) and studying TIL immune properties ex vivo. After we studied ACT for more than thirty years, we are now going to study T-cells' immune properties for personalized immunotherapy. A spatial-quantitative network model can be applied to analyze the immune properties of T-cells and then develop new strategies under a machine learning algorithm. This study utilized STAT3 complex down-regulation for IL-2 inducing cytotoxic T-lymphocyte (CTL) to study T-cell characteristics. After studying spatial-order regulation from the SMAD-STAT3 axis of multiple protein complexes, we focused on a second protein, or STAT3, knockdown for IL-2-inducing CTL. Because machine learning algorithms require a good pathway configuration to mine targeting genes and targeting drugs, the spatial-time quantification network can address the spatial competition of complex proteins binding to downstream proteins/DNA in network analysis. The research demonstrated that STAT3 downregulation can reactivate CTL under a spatial-order quantification network combined with IL2 inducing. After studying the algorithm of a spatial quantitative network under STAT3 downregulation, we can use the spatial-complex configurations to analyze complex regulation in artificial intelligence for future personalized therapy.
Citation: Yun Yang, Shuyi Chen, Wenqian Li, Jason Chengwu Duan, Yuan Yuan, Victor S. Xie, Biaoru Li. STAT3 down-regulation for IL-2 inducing TIL-A clinical clue for spatial quantitative pathway analysis[J]. AIMS Allergy and Immunology, 2024, 8(4): 283-295. doi: 10.3934/Allergy.2024017
Tumor-infiltrating lymphocytes (TILs) can be used in two clinical areas: Adoptive cell therapy (ACT) and studying TIL immune properties ex vivo. After we studied ACT for more than thirty years, we are now going to study T-cells' immune properties for personalized immunotherapy. A spatial-quantitative network model can be applied to analyze the immune properties of T-cells and then develop new strategies under a machine learning algorithm. This study utilized STAT3 complex down-regulation for IL-2 inducing cytotoxic T-lymphocyte (CTL) to study T-cell characteristics. After studying spatial-order regulation from the SMAD-STAT3 axis of multiple protein complexes, we focused on a second protein, or STAT3, knockdown for IL-2-inducing CTL. Because machine learning algorithms require a good pathway configuration to mine targeting genes and targeting drugs, the spatial-time quantification network can address the spatial competition of complex proteins binding to downstream proteins/DNA in network analysis. The research demonstrated that STAT3 downregulation can reactivate CTL under a spatial-order quantification network combined with IL2 inducing. After studying the algorithm of a spatial quantitative network under STAT3 downregulation, we can use the spatial-complex configurations to analyze complex regulation in artificial intelligence for future personalized therapy.

| [1] | Li BR, Tong SQ, Zhang XH, et al. (1994) A new experimental and clinical approach of combining usage of highly active tumor-infiltrating lymphocytes and highly sensitive antitumor drugs for the advanced malignant tumor. Chin Med J (Engl) 107: 803-807. |
| [2] |
Li B, Tong S, Zhang XH, et al. (2020) Chapter-9: Development of adoptive T-cell immunotherapy-Future of personalized immunotherapy. Personalized immunotherapy for tumor diseases and beyond . UAE: Bentham Science Publishers 137-154. https://doi.org/10.2174/9789811482755120010012 
|
| [3] | Hu BC, Li GW, Cheng W, et al. (1997) Clinical application of infiltrating lymphocytes in malignant brain tumors. J Immunol (Chin) 2: 1-2. |
| [4] | Hua ZD, Lu J, Li HF, et al. (1996) Clinical study of tumor infiltrating lymphocytes in ovarian cancer. Chin J Obstet Gynecol 31: 55-57. |
| [5] | Lu J, Hu LLW, Hua ZD, et al. (1996) Analysis of the therapeutic effects of different therapeutic approaches for TIL. Chin J Cancer Biother 3: 127-129. |
| [6] | Cai XM, Lu J, Hua ZD, et al. (1996) Clinical application of TIL from different sources. J Immunol (Chin) 12: 251-254. |
| [7] | Li B, Ding J, Larson A, et al. (1999) Tumor tissue recycling-a new combination treatment for solid tumors: Experimental and preliminarily clinical research. In Vivo 13: 433-438. |
| [8] |
Chen HY, Huang N, Tian HW, et al. (2021) Splenectomy provides protective effects against CLP-induced sepsis by reducing TRegs and PD-1/PD-L1 expression. Int J Biochem Cell Biolo 136: 105970. https://doi.org/10.1016/j.biocel.2021.105970
|
| [9] |
Wu L, Mao L, Liu JF, et al. (2019) Blockade of TIGIT/CD155 signaling reverses T-cell exhaustion and enhances antitumor capability in head and neck squamous cell carcinoma. Cancer Immunol Res 7: 1700-1713. https://doi.org/10.1158/2326-6066.CIR-18-0725
|
| [10] |
Labiano S, Palazon A, Melero I (2015) Immune response regulation in the tumor microenvironment by hypoxia. Semin Oncol 42: 378-386. https://doi.org/10.1053/j.seminoncol.2015.02.009
|
| [11] |
Chevolet I, Speeckaert R, Schreuer M, et al. (2015) Clinical significance of plasmacytoid dendritic cells and myeloid-derived suppressor cells in melanoma. J Transl Med 13: 1-9. https://doi.org/10.1186/s12967-014-0376-x
|
| [12] |
Minnema-Luiting J, Vroman H, Aerts J, et al. (2018) Heterogeneity in immune cell content in malignant pleural mesothelioma. Int J Mol Sci 19: 1041. https://doi.org/10.3390/ijms19041041
|
| [13] |
Oweida AJ, Darragh L, Phan A, et al. (2019) STAT3 modulation of regulatory T cells in response to radiation therapy in head and neck cancer. J Natl Cancer Inst 111: 1339-1349. https://doi.org/10.1093/jnci/djz036
|
| [14] |
Dong S, Guo XG, Han F, et al. (2022) Emerging role of natural products in cancer immunotherapy. Acta Pharm Sin B 12: 1163-1185. https://doi.org/10.1016/j.apsb.2021.08.020
|
| [15] |
Liao F, Liu L, Luo E, et al. (2018) Curcumin enhances anti-tumor immune response in tongue squamous cell carcinoma. Arch Oral Biol 92: 32-37. https://doi.org/10.1016/j.archoralbio.2018.04.015
|
| [16] |
Zhai NC, Li HJ, Song HX, et al. (2017) Hepatitis C virus induces MDSCs-like monocytes through TLR2/PI3K/AKT/STAT3 signaling. PLoS One 12: e0170516. https://doi.org/10.1371/journal.pone.0170516
|
| [17] |
Huang LL, Zhao Y, Shan MY, et al. (2023) Targeting crosstalk of STAT3 between tumor-associated M2 macrophages and Tregs in colorectal cancer. Cancer Biol Ther 24: 2226418. https://doi.org/10.1080/15384047.2023.2226418
|
| [18] |
Xiong WJ, Chen YH, Zhang CT, et al. (2023) Pharmacologic inhibition of IL11/STAT3 signaling increases MHC-I expression and T cell infiltration. J Transl Med 21: 416. https://doi.org/10.1186/s12967-023-04079-6
|
| [19] |
Yu L, Ji Kim H, Kim B, et al. (2023) Ethacrynic acid suppresses B7-H4 expression involved in epithelial-mesenchymal transition of lung adenocarcinoma cells via inhibiting STAT3 pathway. Biochem Pharmacol 212: 115537. https://doi.org/10.1016/j.bcp.2023.115537
|
| [20] |
Alahdal M, Xing Y, Tang TT, et al. (2018) 1-Methyl-D-tryptophan reduces tumor CD133+ cells, Wnt/β-catenin and NF-κβp65 while enhances lymphocytes NF-κβ2, STAT3, and STAT4 pathways in murine pancreatic adenocarcinoma. Sci Rep 8: 9869. https://doi.org/10.1038/s41598-018-28238-8
|
| [21] |
Han QJ, Wang YQ, Pang M, et al. (2017) STAT3-blocked whole-cell hepatoma vaccine induces cellular and humoral immune response against HCC. J Exp Clin Cancer Res 36: 1-11. https://doi.org/10.1186/s13046-017-0623-0
|
| [22] |
Li M, Shang DS, Zhao WD, et al. (2009) Amyloid β interaction with receptor for advanced glycation end products up-regulates brain endothelial CCR5 expression and promotes T cells crossing the blood-brain barrier. J Immunol 182: 5778-5788. https://doi.org/10.4049/jimmunol.0803013
|
| [23] | Li B (2008) A strategy to identify genomic expression profiles at single-T-cell level and a small number of cells (review paper). J Biotechnol 7: 71-82. http://dx.doi.org/10.2225/vol8-issue1-fulltext-3 |
| [24] |
Li B, Perabekam S, Liu G, et al. (2002) Experimental and bioinformatics comparison of gene expression between T cells from TIL of liver cancer and T cells from UniGene. J Gastroenterol 37: 275-282. https://doi.org/10.1007/s005350200035
|
| [25] |
Li B, Chang TF, Larson A, et al. (2000) Identification of mRNAs expressed in tumor-infiltrating lymphocytes by a strategy for rapid and high throughput screening. Gene 255: 273-279. https://doi.org/10.1016/S0378-1119(00)00330-9
|
| [26] | Li B (2015) Breakthrough of 2015-Personalized immunotherapy based on individual GWAS and biomarkers. Biomark J 1: 1-2. |
| [27] |
Zhang W, Yan Q, Lin MH, et al. (2020) Immune cells signaling-personalized immunotherapy for tumor diseases and beyond. Personalized Immunotherapy for Tumor Diseases and Beyond . UAE: Bentham Science Publishers 20-42. https://doi.org/10.2174/9789811482755120010005
|
| [28] |
Li B, Zhu XG, Ward CM, et al. (2019) MIR-144-mediated NRF2 gene silencing inhibits fetal hemoglobin expression in sickle cell disease. Exp Hematol 70: 85-96.e5. https://doi.org/10.1016/j.exphem.2018.11.002
|
| [29] |
Ren YX, Yang J, Sun RM, et al. (2016) Viral IL-10 down-regulates the “MHC-I antigen processing operon” through the NF-κB signaling pathway in nasopharyngeal carcinoma cells. Cytotechnology 68: 2625-2636. https://doi.org/10.1007/s10616-016-9987-9
|
| [30] |
Li Z, Abraham BJ, Berezovskaya A, et al. (2017) APOBEC signature mutation generates an oncogenic enhancer that drives LMO1 expression in T-ALL. Leukemia 31: 2057-2064. https://doi.org/10.1038/leu.2017.75
|
| [31] |
Zhang W, Ding JQ, Qu Y, et al. (2009) Genomic expression analysis of quiescent CD8 T-cells from tumor-infiltrating lymphocytes of in vivo liver tumor by single-cell mRNA differential display. Immunology 127: 83-90. https://doi.org/10.1111/j.1365-2567.2008.02926.x
|
| [32] | Li B, Hu HL, Ding J, et al. (2015) Functional cell-proliferation and differentiation by system modeling for cell therapy. Int J Latest Res Sci Technol 4: 180-187. |
| [33] | Li B, Liu G, Hu HL, et al. (2015) Biomarkers analysis for heterogeneous immune responses of quiescent CD8+ cells-a clue for personalized immunotherapy. Biomark J 1: 1-12. |
| [34] |
Min Tao, Li B, Nayini J, et al. (2000) SCF, IL-1 beta, IL-1ra and GM-CSF in the bone marrow and serum of normal individuals and of AML CML patients. Cytokine 12: 699-707. https://doi.org/10.1006/cyto.2000.0666
|
| [35] | Preisler HD, Li B, Yang BL, et al. (2000) Suppression of telomerase activity and cytokine messenger RNA levels in acute myelogenous leukemia cells in vivo in patients by amifostine and interleukin 4. Clin Cancer Res 6: 807-812. |
| [36] |
Mundle SD, Mativi BY, Cartlidge JD, et al. (2000) Signal antonymy unique to myelodysplastic marrows correlates with altered expression of E2F1. Br J Haematol 109: 376-381. https://doi.org/10.1046/j.1365-2141.2000.01937.x
|
| [37] |
Yang J, Li BR, Nayini J, et al. (1999) Tyrosine phosphorylation of Shc proteins in normal CD34+ progenitor cells and leukemia cells. Blood 94: 373-374. https://doi.org/10.1182/blood.V94.1.373.413a48b_373_374
|
| [38] |
Preisler HD, Gao XZ, Ming T, et al. (1999) Marrow cytokine transcripts and the secondary hematologic disorders. Leuk Lymphoma 35: 297-302. https://doi.org/10.3109/10428199909145733
|
| [39] |
Perambakam S, Li B, Preisler H (2001) Quantitation of interferon regulatory factor transcripts in patients with AML. Cancer Invest 19: 346-351.
|
| [40] | Li B (2008) A strategy to identify genomic expression profiles at single-T-cell level and a small number of cells. Electron J Biotechnol 8. http://dx.doi.org/10.2225/vol8-issue1-fulltext-3 |
| [41] | Li B, Zhu YM, Hu BY, et al. (1994) Establishment of a method for separation of tumor infiltrating lymphocytes with high vitality. J Immunol (Chin) 10: 44-47. |
| [42] |
Li S, Perabekam S, Devemy E, et al. (2020) Genetically modified T-cells affinity to tumor cells-development of adoptive T-cell immunotherapy. Personalized Immunotherapy for Tumor Diseases and Beyond . UAE: Bentham Science Publishers 174-149. https://doi.org/10.2174/9789811482755120010014
|
| [43] |
Li B, Xu YB (2022) Single-cell whole RNA sequencing from tumor-infiltrating lymphocyte. New Horizons in Medicine and Medical Research : 167-173. https://doi.org/10.9734/bpi/nhmmr/v10/6129F
|
| [44] |
Liu G, Zheng J, Li B (2020) Bioinformatics of T-cell and primary tumor cells. Personalized Immunotherapy for Tumor Diseases and Beyond . UAE: Bentham Science Publishers 118-136. https://doi.org/10.2174/9789811482755120010011
|
| [45] |
Wang ZH, Hu HL, Zheng J, et al. (2011) Gene expression and pathway analysis of quiescent CD8+ T cells from liver cancer, liver sinusoid and peripheral blood-study on toxicogenomic and prevention targeting. “2011 IEEE 11th International Conference on Bioinformatics and Bioengineering” : 72-77. https://doi.org/10.1109/BIBE.2011.18
|
| [46] |
Li B, Liu G, Zheng J (2020) System modeling of T-cell function development of adoptive T-cell immunotherapy. Personalized Immunotherapy for Tumor Diseases and Beyond . UAE: Bentham Science Publishers 197-223. https://doi.org/10.2174/9789811482755120010015
|
| [47] |
Tan SZ, Zhu X (2020) Molecular targeting checkpoint in cancer. Personalized Immunotherapy for Tumor Diseases and Beyond . UAE: Bentham Science Publishers 70-89. https://doi.org/10.2174/9789811482755120010008
|
| [48] | Hu HL Zhang QH, Li S, et al. (2015) A therapeutic targeting identification from microarray data and quantitative network analysis. Open Access J Sci Technol 3. https://doi.org/10.11131/2015/101114 |
| [49] | Xiao HS, Gao Y, Li S, et al. (2016) A case report of personalized chemotherapy for metastatic cardiac sarcoma. J Clin Epigenet 1: 1-6. |
| [50] | Lu J, Cheng SJ, Li B (2007) Pathway-based approaches for analysis of genome-wide association studies-a case report for metastatic small cell lung cancer. Int J Hematol Ther 10: 2381-1404. |
| [51] |
Li B, Larson A, Li S (2020) Personalized Immunotherapy for Tumor Diseases and Beyond. UAE: Bentham Science Publishers. https://doi.org/10.2174/97898114827551200101
|
| [52] | Ding JQ, Qian GX, Li B, et al. (1995) A preliminary study of tumor necrosis factor gene transduction of tumor infiltrating lymphocytes Application. Chin J Cancer Biother 1: 11-15. |
| [53] |
Jing LH, Li B (2020) Immunoassay of personalized immunotherapy. Personalized Immunotherapy for Tumor Diseases and Beyond . UAE: Bentham Science Publishers 43-56. https://doi.org/10.2174/9789811482755120010006
|
| [54] | Xu YB, Hu HL, Zheng J, et al. (2013) Feasibility of whole RNA sequencing from single-cell mRNA amplification. Genet Res Int . https://doi.org/10.1155/2013/724124 |
| [55] | Li B, Hu HL, Ding JQ, et al. (2015) Functional cell-proliferation and differentiation by system modelling for cell therapy. Int J Latest Res Sci Technol 4: 180-187. Available from: https://www.mnkjournals.com/journal/ijlrst/index.php |
| [56] | Ying XN, Li B (2022) Machine-learning modeling for personalized immunotherapy-an evaluation module. Biomed J Scie Tech Res 47: 38211. https://doi.org/10.26717/BJSTR.2022.47.007462 |
| [57] |
Takeda K, Clausen BE, Kaisho T, et al. (1999) Enhanced Th1 activity and development of chronic enterocolitis in mice devoid of Stat3 in macrophages and neutrophils. Immunity 10: 39-49. https://doi.org/10.1016/S1074-7613(00)80005-9
|
| [58] |
Goswami K, Barik S, Sarkar M, et al. (2014) Targeting STAT3 phosphorylation by neem leaf glycoprotein prevents immune evasion exerted by supraglottic laryngeal tumor induced M2 macrophages. Mol Immunol 59: 119-127. https://doi.org/10.1016/j.molimm.2014.01.015
|
| [59] |
Üremiş MM, Üremis N, Türköz Y (2023) Cucurbitacin E shows synergistic effect with sorafenib by inducing apoptosis in hepatocellular carcinoma cells and regulates Jak/Stat3, ERK/MAPK, PI3K/Akt/mTOR signaling pathways. Steroids 198: 109261. https://doi.org/10.1016/j.steroids.2023.109261
|
| [60] |
Wang G, Yu Y, Sun C, et al. (2016) STAT3 selectively interacts with Smad3 to antagonize TGF-β signaling. Oncogene 35: 4388-4398. https://doi.org/10.1038/onc.2015.446
|
| [61] |
Németh Á, Mózes MM, Calvier L, et al. (2019) The PPARγ agonist pioglitazone prevents TGF-β induced renal fibrosis by repressing EGR-1 and STAT3. BMC Nephrol 20. https://doi.org/10.1186/s12882-019-1431-x
|
| [62] |
Sun ZJ, Lin DD, Shen Y, et al. (2023) Critical role of MXRA7 in differentiation blockade in human acute promyelocytic leukemia cells. Exp Hematol 125: 45-54. https://doi.org/10.1016/j.exphem.2023.07.001
|
| [63] |
Zhu YZ, Wang XH, He ZW, et al. (2023) Enterovirus 71 enters human brain microvascular endothelial cells through an ARF6-mediated endocytic pathway. J Med Virol 95: e28915. https://doi.org/10.1002/jmv.28915
|
| [64] |
Kongkaew T, Thaiwong R, Tudsamran S, et al. (2022) TIL expansion with high dose IL-2 or low dose IL-2 with anti-CD3/anti-CD28 stimulation provides different quality of TIL-expanded T cell clones. J Immunol Methods 503: 113229. https://doi.org/10.1016/j.jim.2022.113229
|
| [65] |
Knochelmann HM, Rivera-Reyes AM, Wyatt MM, et al. (2021) Modeling ex vivo tumor-infiltrating lymphocyte expansion from established solid malignancies. Oncoimmunology 10: 1959101. https://doi.org/10.1080/2162402X.2021.1959101
|
| [66] | Li B (2022) Why will TIL produce different efficacy to treat solid tumor. Front Immunol 2: 1-13. https://doi.org/10.3389/fimmu.2022.973881 |
 allergy-08-04-017-s001.xlsx allergy-08-04-017-s001.xlsx |
 |


Figures(3) / Tables(1)

Yun Yang, Shuyi Chen, Wenqian Li, Jason Chengwu Duan, Yuan Yuan, Victor S. Xie, Biaoru Li. STAT3 down-regulation for IL-2 inducing TIL-A clinical clue for spatial quantitative pathway analysis[J]. AIMS Allergy and Immunology, 2024, 8(4): 283-295. doi: 10.3934/Allergy.2024017







 DownLoad:
DownLoad: