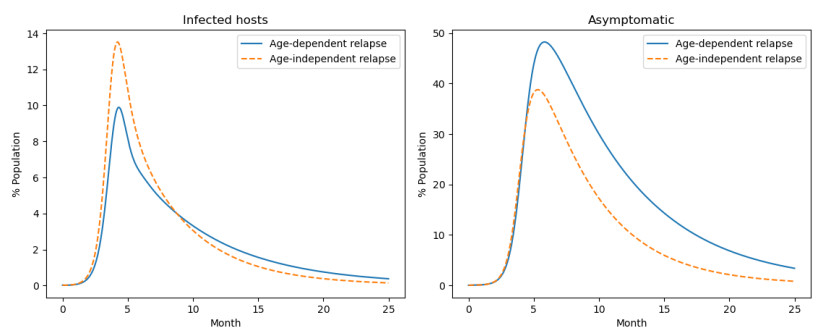
Mosquito-borne infectious diseases represent a significant public health issue. Age has been identified as a key risk factor for these diseases, and another phenomenon reported is relapse, which involves the reappearance of symptoms after a symptom-free period. Recent research indicates that susceptibility to and relapse of mosquito-borne diseases are frequently age-dependent. This paper proposes a new model to better capture the dynamics of mosquito-borne diseases by integrating two age-dependent factors: chronological age and asymptomatic-infection age. Chronological age refers to the time elapsed from the date of birth of the host to the present time. On the other hand, asymptomatic infection age denotes the time elapsed since the host became asymptomatic after the primary infection. The system of integro-differential equations uses flexible, unspecified functions to represent these dependencies, assuming they are integrable. We analyzed the global stability of both the disease-free and endemic equilibrium states using the direct Lyapunov method with Volterra-type Lyapunov functionals. Additionally, the paper explores several special cases involving well-known host-vector models.
Citation: Maria Guadalupe Vazquez-Peña, Cruz Vargas-De-León, Jorge Velázquez-Castro. Global stability for a mosquito-borne disease model with continuous-time age structure in the susceptible and relapsed host classes[J]. Mathematical Biosciences and Engineering, 2024, 21(11): 7582-7600. doi: 10.3934/mbe.2024333
Mosquito-borne infectious diseases represent a significant public health issue. Age has been identified as a key risk factor for these diseases, and another phenomenon reported is relapse, which involves the reappearance of symptoms after a symptom-free period. Recent research indicates that susceptibility to and relapse of mosquito-borne diseases are frequently age-dependent. This paper proposes a new model to better capture the dynamics of mosquito-borne diseases by integrating two age-dependent factors: chronological age and asymptomatic-infection age. Chronological age refers to the time elapsed from the date of birth of the host to the present time. On the other hand, asymptomatic infection age denotes the time elapsed since the host became asymptomatic after the primary infection. The system of integro-differential equations uses flexible, unspecified functions to represent these dependencies, assuming they are integrable. We analyzed the global stability of both the disease-free and endemic equilibrium states using the direct Lyapunov method with Volterra-type Lyapunov functionals. Additionally, the paper explores several special cases involving well-known host-vector models.

| [1] |
B. Chala, F. Hamde, Emerging and re-emerging vector-borne infectious diseases and the challenges for control: a review, Front. Public Health, 9 (2021), 715759. https://doi.org/10.3389/fpubh.2021.715759 doi: 10.3389/fpubh.2021.715759

|
| [2] |
R. T. Jones, T. H. Ant, M. M. Cameron, J. G. Logan, Novel control strategies for mosquito-borne diseases, Phil. Trans. R. Soc. B, 376 (2021), 20190802. https://doi.org/10.1098/rstb.2019.0802 doi: 10.1098/rstb.2019.0802
|
| [3] |
H. Hasyim, W. C. Dewi, W. C. Lestari, R. Flora, N. Novrikasari, I. A. Liberty, et al., Risk factors of malaria transmission in mining workers in Muara Enim, South Sumatra, Indonesia, Sci. Rep., 13 (2023), 14755. https://doi.org/10.1038/s41598-023-40418-9 doi: 10.1038/s41598-023-40418-9
|
| [4] |
N. Thang, C. Duy, C. Xuan, H. Thi, H. Mats, N. Hung, et al., Risk factors of dengue fever in an urban area in Vietnam: a case-control study, BMC Public Health, 21 (2021), 664. https://doi.org/10.1186/s12889-021-10687-y doi: 10.1186/s12889-021-10687-y
|
| [5] |
L. M. Verhagen, R. de Groot, Dengue in children, J. Infect., 69 (2014), S77–S86. https://doi.org/10.1016/j.jinf.2014.07.020 doi: 10.1016/j.jinf.2014.07.020
|
| [6] |
I. C. Ster, A. Rodriguez, N. C. Romero, N. C. Lopez, M. Chico, J. Montgomery, et al., Age-dependent seroprevalence of dengue and chikungunya: inference from a cross-sectional analysis in Esmeraldas Province in coastal Ecuador, BMJ Open, 10 (2020), e040735. https://doi.org/10.1136/bmjopen-2020-040735 doi: 10.1136/bmjopen-2020-040735
|
| [7] |
B. Nadjm, R. H. Behrens, Malaria: An update for physicians, Infect. Dis. Clin., 26 (2012), 243–259. https://doi.org/10.1016/j.idc.2012.03.010 doi: 10.1016/j.idc.2012.03.010
|
| [8] |
A. K. Supriatna, E. Soewono, S. A. van Gils, A two-age-classes dengue transmission model, Math. Biosci., 216 (2008), 114–121. https://doi.org/10.1016/j.mbs.2008.08.011 doi: 10.1016/j.mbs.2008.08.011
|
| [9] |
E. Couturier, F. Guillemin, M. Mura, L. León, J. M. Virion, J. M. Letort, et al., Impaired quality of life after Chikungunya virus infection: a 2-year follow-up study, Rheumatology, 51 (2012), 1315–1322. https://doi.org/10.1093/rheumatology/kes015 doi: 10.1093/rheumatology/kes015
|
| [10] |
I. X. De La Cruz-Castro, E. Nava-Aguilera, A. Morales-Pérez, Á. F. Betanzos-Reyes, M. Flores-Moreno, L. Morales-Nava, et al., Factors associated with Chikungunya relapse in Acapulco, México, Vector-Borne Zoonotic Dis, 20 (2020), 782–787. https://doi.org/10.1089/vbz.2020.2615 doi: 10.1089/vbz.2020.2615
|
| [11] |
D. Aldila, T. Götz, T. Soewono, An optimal control problem arising from a dengue disease transmission model, Math. Biosci., 242 (2013), 9–16. https://doi.org/10.1016/j.mbs.2012.11.014 doi: 10.1016/j.mbs.2012.11.014
|
| [12] |
E. Beretta, V. Capasso, D. G. Garao, A mathematical model for malaria transmission with asymptomatic carriers and two age groups in the human population, Math. Biosci., 300 (2018), 87–101. https://doi.org/10.1016/j.mbs.2018.03.024 doi: 10.1016/j.mbs.2018.03.024
|
| [13] |
A. Li-Martín, R. Reyes-Carreto, C. Vargas-De-León, Dynamics of a dengue disease transmission model with two-stage structure in the human population, Math. Biosci. Eng., 20 (2023), 955–974. https://doi.org/10.3934/mbe.2023044 doi: 10.3934/mbe.2023044
|
| [14] |
A. Chamnan, P. Pongsumpun, I. M. Tang, N. Wongvanich, Effect of a vaccination against the dengue fever epidemic in an age structure population: From the perspective of the local and global stability analysis, Mathematics, 10 (2022), 904. https://doi.org/10.3390/math10060904 doi: 10.3390/math10060904
|
| [15] |
H. F. Huo, G. M. Qiu, Stability of a mathematical model of malaria transmission with relapse, Abstr. Appl. Anal., 1 (2014), 289349. https://doi.org/10.1155/2014/289349 doi: 10.1155/2014/289349
|
| [16] |
G. M. Ghosh, S. Olaniyi, O. S. Obabiyi, Mathematical analysis of reinfection and relapse in malaria dynamics, Appl. Math. Comput., 373 (2020), 125044. https://doi.org/10.1016/j.amc.2020.125044 doi: 10.1016/j.amc.2020.125044
|
| [17] |
O. S. Vázquez-Peña, C. Vargas-De-León, J. F. Camacho-Pérez, J. Velázquez-Castro, Analysis and Bayesian estimation of a model for Chikungunya dynamics with relapse: An outbreak in Acapulco, Mexico, Math. Biosci. Eng., 20 (2023), 18123–18145. https://doi.org/10.3934/mbe.2023805 doi: 10.3934/mbe.2023805
|
| [18] |
C. Vargas-De-León, L. Esteva, A. Korobeinikov, Age-dependency in host-vector models: The global analysis, Appl. Math. Comput., 243 (2014), 969–981. http://dx.doi.org/10.1016/j.amc.2014.06.042 doi: 10.1016/j.amc.2014.06.042
|
| [19] |
C. Vargas-De-León, Global stability properties of age‐dependent epidemic models with varying rates of recurrence, Math. Methods Appl. Sci., 39 (2016), 2057–2064. https://doi.org/10.1002/mma.3621 doi: 10.1002/mma.3621
|
| [20] | H. L. Smith, H. L. Thieme, Dynamical Systems and Population Persistence, American Mathematical Society, 2011. |
| [21] |
L. Esteva, C. Vargas, Analysis of a dengue disease transmission model, Math. Biosci., 150 (1998), 131–151. https://doi.org/10.1016/S0025-5564(98)10003-2 doi: 10.1016/S0025-5564(98)10003-2
|
| [22] | C. Hirsch, Numerical Computation of Internal and External Flows: Fundamentals of Computational Fluid Dynamics, Elsevier/Butterworth-Heinemann, 2007. |
| [23] |
G. Cruz-Pacheco, L. Esteva, C. Vargas, Vaccination strategies for SIR vector-transmitted diseases, Bull. Math. Biol., 76 (2014), 2073–2090. https://doi.org/10.1007/s11538-014-9999-6 doi: 10.1007/s11538-014-9999-6
|
| [24] |
M. M. Haby, M. Pinart, V. Elias, L. Reveiz, Prevalence of asymptomatic Zika virus infection: a systematic review, Bull. World Health Organ., 96 (2018), 402–413D. https://doi.org/10.2471/BLT.17.201541 doi: 10.2471/BLT.17.201541
|
| [25] |
A. E. Ades, A. Soriano-Arandes, A. Alarcon, F. Bonfante, C. Thorne, C. S. Peckham, et al., Vertical transmission of Zika virus and its outcomes: a Bayesian synthesis of prospective studies, Lancet Infect. Dis., 21 (2021), 537–545. https://doi.org/10.1016/s1473-3099(20)30432-1 doi: 10.1016/s1473-3099(20)30432-1
|

Figures(2) / Tables(2)

Maria Guadalupe Vazquez-Peña, Cruz Vargas-De-León, Jorge Velázquez-Castro. Global stability for a mosquito-borne disease model with continuous-time age structure in the susceptible and relapsed host classes[J]. Mathematical Biosciences and Engineering, 2024, 21(11): 7582-7600. doi: 10.3934/mbe.2024333







 DownLoad:
DownLoad: