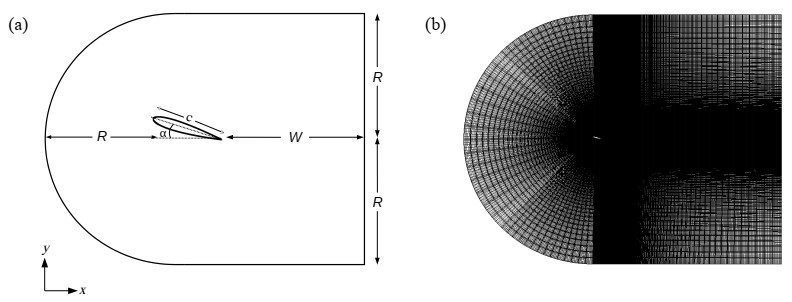
A high-order spectral element method was used to perform three-dimensional direct numerical simulations of the flow past a NACA0012 airfoil. We considered a Reynolds number $ Re = 1000 $ and two different angles of attack, $ \alpha = 11^\circ $ and $ \alpha = 16^\circ $, to study the two- to three-dimensional wake transition. A boundary layer separation was observed for both angles of attack with the separation point closer to the leading edge for $ \alpha = 16^\circ $. The downstream of the airfoil exhibited streamwise vortical structures formed in the braid regions connecting the primary vortices for $ \alpha = 16^\circ $, while only shed vortices were observed for $ \alpha = 11^\circ $. The formation of these streamwise structures were explained by the presence of a reverse flow from the lower surface for $ \alpha = 16^\circ $, enhancing shearing effects. The early-stage development of the three-dimensional wake, in the case of $ \alpha = 16^\circ $, was characterized by the formation of a spanwise sinusoidal velocity whose amplitude increased exponentially over time. The flow on the upper surface experienced a higher strain field which pulled up small disturbances from the airfoil surface and formed regions of concentrated vortical structures. These structures were subjected to stretching under the strain field and later advected downstream of the airfoil.
Citation: Hussein Kokash, G. Gilou Agbaglah. A two- to three-dimensional wake transition mechanism induced by the angle of attack of a NACA0012 airfoil[J]. Metascience in Aerospace, 2024, 1(3): 329-345. doi: 10.3934/mina.2024015
A high-order spectral element method was used to perform three-dimensional direct numerical simulations of the flow past a NACA0012 airfoil. We considered a Reynolds number $ Re = 1000 $ and two different angles of attack, $ \alpha = 11^\circ $ and $ \alpha = 16^\circ $, to study the two- to three-dimensional wake transition. A boundary layer separation was observed for both angles of attack with the separation point closer to the leading edge for $ \alpha = 16^\circ $. The downstream of the airfoil exhibited streamwise vortical structures formed in the braid regions connecting the primary vortices for $ \alpha = 16^\circ $, while only shed vortices were observed for $ \alpha = 11^\circ $. The formation of these streamwise structures were explained by the presence of a reverse flow from the lower surface for $ \alpha = 16^\circ $, enhancing shearing effects. The early-stage development of the three-dimensional wake, in the case of $ \alpha = 16^\circ $, was characterized by the formation of a spanwise sinusoidal velocity whose amplitude increased exponentially over time. The flow on the upper surface experienced a higher strain field which pulled up small disturbances from the airfoil surface and formed regions of concentrated vortical structures. These structures were subjected to stretching under the strain field and later advected downstream of the airfoil.

| [1] |
Romeo G, Frulla G, Cestino E, et al. (2004) HELIPLAT: Design, aerodynamic, structural analysis of long-endurance solar-powered stratospheric platform. J Aircraft 41: 1505–1520. https://doi.org/10.2514/1.2723 doi: 10.2514/1.2723

|
| [2] |
Hassanalian M, Abdelkefi A (2017) Classifications, applications, and design challenges of drones: A review. Prog Aerosp Sci 91: 99–131.https://doi.org/10.1016/j.paerosci.2017.04.003 doi: 10.1016/j.paerosci.2017.04.003
|
| [3] | Carmichael BH (1981) Low-Reynolds number airfoil survey. 165803, NASA. |
| [4] |
Lin YF, Lam K, Zou L, et al. (2013) Numerical study of flows past airfoils with wavy surfaces. J Fluid Struct 36: 136–148. https://doi.org/10.1016/j.jfluidstructs.2012.09.008 doi: 10.1016/j.jfluidstructs.2012.09.008
|
| [5] |
Zhang W, Cheng W, Gao W, et al. (2015) Geometrical effects on the airfoil flow separation and transition. Comput Fluids 116: 60–73. https://doi.org/10.1016/j.compfluid.2015.04.014 doi: 10.1016/j.compfluid.2015.04.014
|
| [6] | Karasu I, Genç MS, Açikel HH (2013) Numerical study on low Reynolds number flows over an Aerofoil. J Appl Mech Eng 2: 1000131. |
| [7] |
Amiralaei MR, Alighanbari H, Hashemi SM (2010) An investigation into the effects of unsteady parameters on the aerodynamics of a low Reynolds number pitching airfoil. J Fluids Struct 26: 979–993. https://doi.org/10.1016/j.jfluidstructs.2010.06.004 doi: 10.1016/j.jfluidstructs.2010.06.004
|
| [8] | Mueller TJ (1999) Aerodynamic Measurements at Low Reynolds Numbers for Fixed Wing Micro-Air Vehicles, in: RTO AVT/VKI Special Course on Development and Operation of UAVs for Military and Civil Applications, Hessert Center for Aerospace Research, University of Notre Dame, VKI, Belgium, 13–17. |
| [9] |
Munday PM, Taira K, Suwa T, et al. (2015) Nonlinear Lift on a Triangular Airfoil in Low-Reynolds-Number Compressible Flow. J Aircraft 52: 924–931. https://doi.org/10.2514/1.C032983 doi: 10.2514/1.C032983
|
| [10] |
Kurtulus DF (2016) On the wake pattern of symmetric airfoils for different incidence angles at Re = 1000. Int J Micro Air Veh 8: 109–139. https://doi.org/10.1177/1756829316653700 doi: 10.1177/1756829316653700
|
| [11] |
Rossi E, Colagrossi A, Oger G, et al. (2018) Multiple bifurcations of the flow over stalled airfoils when changing the Reynolds number. J Fluid Mech 846: 356–391. https://doi.org/10.1017/jfm.2018.189 doi: 10.1017/jfm.2018.189
|
| [12] |
Genç MS, Karasu İ, Açıkel HH (2012) An experimental study on aerodynamics of NACA2415 aerofoil at low Re numbers. Exp Therm Fluid Sci 39: 252–264. https://doi.org/10.1016/j.expthermflusci.2012.01.029 doi: 10.1016/j.expthermflusci.2012.01.029
|
| [13] |
Ducoin A, Loiseau JC, Robinet JC (2016) Numerical investigation of the interaction between laminar to turbulent transition and the wake of an airfoil. Eur J Mech B-Fluid 57: 231–248. https://doi.org/10.1016/j.euromechflu.2016.01.005 doi: 10.1016/j.euromechflu.2016.01.005
|
| [14] | Shelton A, Abras J, Hathaway B, et al. (2005) An investigation of the numerical prediction of static and dynamic stall. in: Proceedings of the 61st American Helicopter Society Annual Forum, 61: 1826. |
| [15] |
Genç MS (2010) Numerical Simulation of Flow over a Thin Aerofoil at a High Reynolds Number Using a Transition Model. P I Mech Eng C-J Mec 224: 2155–2164. https://doi.org/10.1243/09544062JMES2121 doi: 10.1243/09544062JMES2121
|
| [16] |
Hain R, Kähler C J, Radespiel R (2019) Dynamics of laminar separation bubbles at low-Reynolds-number aerofoils. J Fluid Mech 630: 129–153. https://doi.org/10.1017/S0022112009006661 doi: 10.1017/S0022112009006661
|
| [17] |
Jones LE, Sandberg RD, Sandham ND (2008) Direct numerical simulations of forced and unforced separation bubbles on an airfoil at incidence. J Fluid Mech 602: 175–207. https://doi.org/10.1017/S0022112008000864 doi: 10.1017/S0022112008000864
|
| [18] |
Zhou Y, Alam MM, Yang HX, et al. (2011) Fluid forces on a very low Reynolds number airfoil and their prediction. Int J Numer Method H 32: 329–339. https://doi.org/10.1016/j.ijheatfluidflow.2010.07.008 doi: 10.1016/j.ijheatfluidflow.2010.07.008
|
| [19] |
Wang S, Zhou Y, Alam MM, et al. (2014) Turbulent intensity and Reynolds number effects on an airfoil at low Reynolds numbers. Phys Fluids 26: 115107. https://doi.org/10.1063/1.4901969 doi: 10.1063/1.4901969
|
| [20] |
Hoarau Y, Braza M, Ventikos Y, et al. (2003) Organized modes and the three-dimensional transition to turbulence in the incompressible flow around a NACA0012 wing. J Fluid Mech 496: 63–72. https://doi.org/10.1017/S0022112003006530 doi: 10.1017/S0022112003006530
|
| [21] |
Huang RF, Wu JY, Jeng JH, et al. (2001) Surface flow and vortex shedding of an impulsively started wing. J Fluid Mech 441: 265–292. https://doi.org/10.1017/S002211200100489X doi: 10.1017/S002211200100489X
|
| [22] | Lissaman PBS (1983) Low-Reynolds-Number Airfoils. Annu Rev Fluid Mech 15: 223–239. |
| [23] | Klose BF, Spedding GR, Jacobs GB (2021) Direct numerical simulation of cambered airfoil aerodynamics at Re = 20,000. arXiv. https://doi.org/10.48550/arXiv.2108.04910 |
| [24] | Williamson CHK (1988) The existence of two stages in the transition to three‐dimensionality of a cylinder wake. Phys Fluids 31: 3165–3168. |
| [25] | Williamson CHK (1996) Vortex dynamics in the cylinder wake. Ann Rev Fluid Mech 28: 477–539. |
| [26] |
Barkley D, Henderson RD (1996) Three-dimensional Floquet stability analysis of the wake of a circular cylinder. J Fluid Mech 322: 215–241. https://doi.org/10.1017/S0022112096002777 doi: 10.1017/S0022112096002777
|
| [27] |
Robichaux J, Balachandar S, Vanka SP (1999) Three-dimensional Floquet instability of the wake of square cylinder. Phys Fluids 11: 560–578. https://doi.org/10.1063/1.869930 doi: 10.1063/1.869930
|
| [28] |
Saha AK, Biswas G, Muralidhar K (2003) Three-dimensional study of flow past a square cylinder at low Reynolds numbers. Int J Heat Fluid Flow 24: 54–66. https://doi.org/10.1016/S0142-727X(02)00208-4 doi: 10.1016/S0142-727X(02)00208-4
|
| [29] |
Luo SC, Tong XH, Khoo BC (2007) Transition phenomena in the wake of a square cylinder. J Fluids Struct 23: 227–248. https://doi.org/10.1016/j.jfluidstructs.2006.08.012 doi: 10.1016/j.jfluidstructs.2006.08.012
|
| [30] |
Luo SC, Chew YT, Ng YT (2003) Characteristics of square cylinder wake transition flows. Phys Fluids 15: 2549–2559. https://doi.org/10.1063/1.1596413 doi: 10.1063/1.1596413
|
| [31] |
Barkley D, Tuckerman LS, Golubitsky M (2000) Bifurcation theory for three-dimensional flow in the wake of a circular cylinder Phys Rev E 61: 5247–5252. https://doi.org/10.1103/PhysRevE.61.5247 doi: 10.1103/PhysRevE.61.5247
|
| [32] |
Blackburn HM, Lopez JM (2003) On three-dimensional quasiperiodic Floquet instabilities of two-dimensional bluff body wakes. Phys Fluids 15: L57–L60. https://doi.org/10.1063/1.1591771 doi: 10.1063/1.1591771
|
| [33] |
Meneghini JR, Carmo BS, Tsiloufas SP, et al. (2011) Wake instability issues: From circular cylinders to stalled airfoils. J Fluids Struct 27: 694–701. https://doi.org/10.1016/j.jfluidstructs.2011.03.018 doi: 10.1016/j.jfluidstructs.2011.03.018
|
| [34] |
Deng J, Sun L, Shao X (2017) Floquet stability analysis in the wake of a NACA0015 airfoil at post-stall angles of attack. Phys Fluids 29: 094104. https://doi.org/10.1063/1.5003578 doi: 10.1063/1.5003578
|
| [35] |
He W, Gioria RS, Pérez JM, et al. (2017) Linear instability of low Reynolds number massively separated flow around three NACA airfoils. J Fluid Mech 811: 701–741. https://doi.org/10.1017/jfm.2016.778 doi: 10.1017/jfm.2016.778
|
| [36] |
Gupta S, Zhao J, Sharma A, et al. (2023) Two- and three-dimensional wake transitions of a NACA0012 airfoil. J Fluid Mech 954: A26. https://doi.org/10.1017/jfm.2022.958 doi: 10.1017/jfm.2022.958
|
| [37] |
Kouser T, Xiong Y, Yang D, et al. (2021) Direct numerical simulations on the three-dimensional wake transition of flows over NACA0012 airfoil at Re = 1000. Int J Micro Air Veh 13: 1–15. https://doi.org/10.1177/17568293211055656 doi: 10.1177/17568293211055656
|
| [38] |
Xia T, Dong H, Yang L, et al. (2021) Investigation on flow structure and aerodynamic characteristics over an airfoil at low Reynolds number–A review. AIP Adv 11: 050701. https://doi.org/10.1063/5.0044717 doi: 10.1063/5.0044717
|
| [39] | Patera AT (1984) A spectral element method for fluid dynamics: Laminar flow in a channel expansion. J Comput Phys 54: 468–488. |
| [40] | Fischer P, Kruse J, Mullen J, et al. Nek5000: Open source spectral element CFD solver. Argonne National Laboratory, Mathematics and Computer Science Division, Argonne, IL, Available from: https://nek5000.mcs.anl.gov/index.php/MainPage2. |
| [41] | Deville MO, Fischer PF, Mund EH (2002) High-Order Methods for Incompressible Fluid Flow, Cambridge Monographs on Applied and Computational Mathematics. Cambridge University Press. |
| [42] | Cummings RM, Morton SA, Mason WH (2015) Applied Computational Aerodynamics, Cambridge Aerospace Series, Cambridge University Press. |
| [43] |
Hoarau Y, Braza M, Ventikos Y, et al. (2006) First stages of the transition to turbulence and control in the incompressible detached flow around a NACA0012 wing. Int J Heat Fluid Fl 27: 878–886. https://doi.org/10.1016/j.ijheatfluidflow.2006.03.026 doi: 10.1016/j.ijheatfluidflow.2006.03.026
|
| [44] |
Uranga A, Persson PO, Drela M, et al. (2011) Implicit Large Eddy Simulation of transition to turbulence at low Reynolds numbers using a discontinuous galerkin method. Int J Numer Meth Eng 87: 232–261. https://doi.org/10.1002/nme.3036 doi: 10.1002/nme.3036
|
| [45] | Khalid MSU, Akhtar I (2012) Characteristics of Flow Past a Symmetric Airfoil at Low Reynolds Number: A Nonlinear Perspective. ASME International Mechanical Engineering Congress and Exposition, American Society of Mechanical Engineers, 167–175. https://doi.org/10.1115/IMECE2012-87389 |
| [46] |
Hongquan Z(1998) Centrifugal instability and the rib vortices in the cylinder wake. Acta Mech Sinica 14: 104–112. https://doi.org/10.1007/BF02487745 doi: 10.1007/BF02487745
|
| [47] |
Agbaglah G, Mavriplis C (2017) Computational analysis of physical mechanisms at the onset of three-dimensionality in the wake of a square cylinder. J Fluid Mech 833: 631–647. https://doi.org/10.1017/jfm.2017.713 doi: 10.1017/jfm.2017.713
|
| [48] |
Agbaglah G, Mavriplis C (2019) Three-dimensional wakes behind cylinders of square and circular cross-section: early and long-time dynamics. J Fluid Mech 870: 419–432. https://doi.org/10.1017/jfm.2019.265 doi: 10.1017/jfm.2019.265
|
| [49] |
Kokash H, Agbaglah GG (2022) On the origin of mode B instability of the wake of a square cylinder. Phys Fluids 34: 074116. https://doi.org/10.1063/5.0101403 doi: 10.1063/5.0101403
|
| [50] |
Kouser T, Xiong Y, Yang D, et al. (2022) Numerical study on drag reduction and wake modification for the flows over a hydrofoil in presence of surface heterogeneity. Adv Mech Eng 14: 1–14. https://doi.org/10.1177/168781402210753 doi: 10.1177/168781402210753
|
| [51] |
Lasheras JC, Choi H (1988) Three-dimensional instability of a plane free shear layer: an experimental study of the formation and evolution of streamwise vortices. J Fluid Mech 189: 53–86. https://doi.org/10.1017/S0022112088000916 doi: 10.1017/S0022112088000916
|
| [52] |
Pierrehumbert RT, Widnall SE (1982) The two- and three-dimensional instabilities of a spatially periodic shear layer. J Fluid Mech 114: 59–59. https://doi.org/10.1017/S0022112082000044 doi: 10.1017/S0022112082000044
|
| [53] |
Tseng CC, Hu HA (2015) Flow dynamics of a pitching foil by eulerian and lagrangian viewpoints. AIAA J 54: 2. https://doi.org/10.2514/1.J053619 doi: 10.2514/1.J053619
|







Figures(8) / Tables(1)
Hussein Kokash, G. Gilou Agbaglah. A two- to three-dimensional wake transition mechanism induced by the angle of attack of a NACA0012 airfoil[J]. Metascience in Aerospace, 2024, 1(3): 329-345. doi: 10.3934/mina.2024015







 DownLoad:
DownLoad: