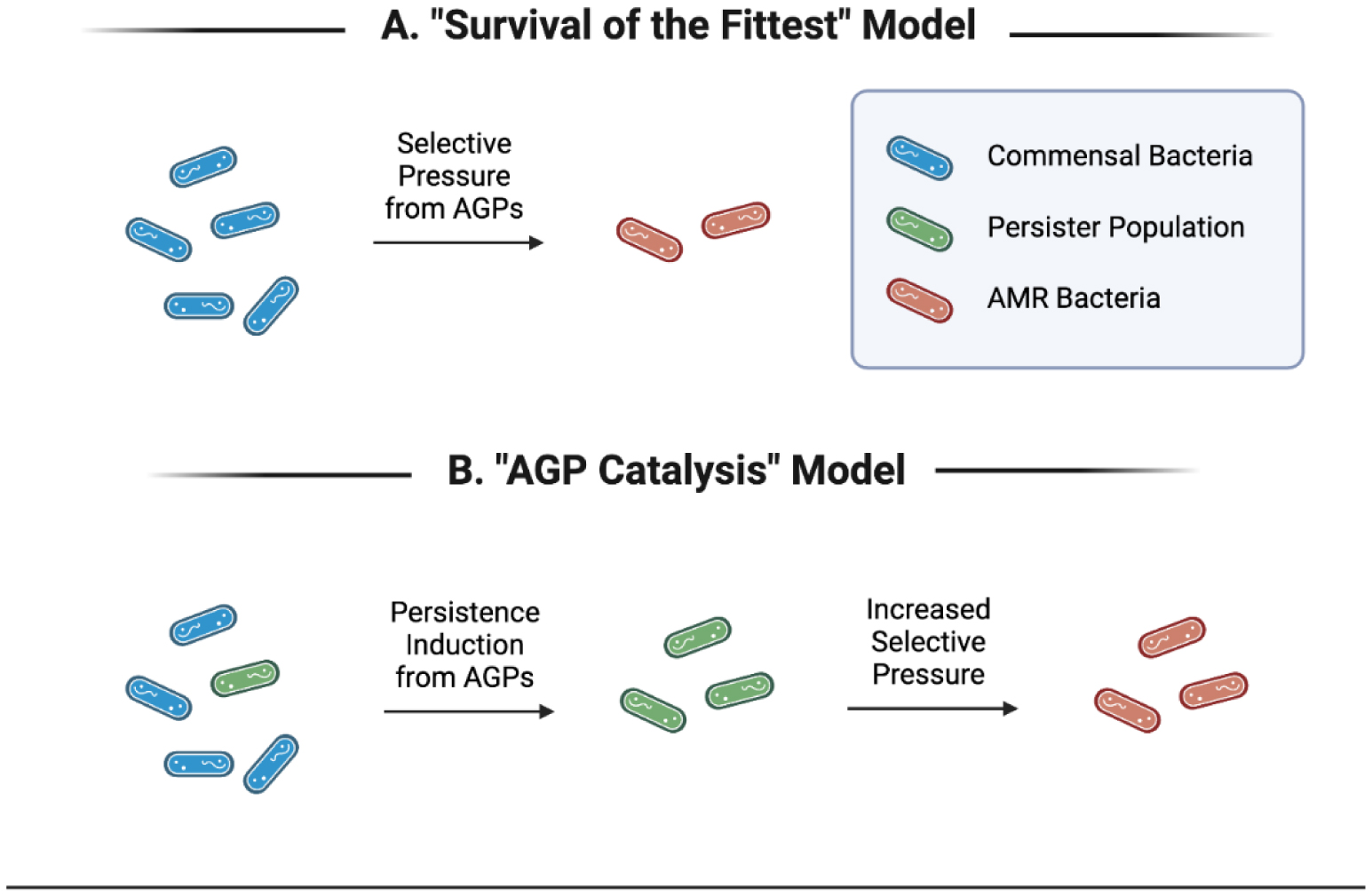
The creation and continued development of antibiotics have revolutionized human health and disease for the past century. The emergence of antimicrobial resistance represents a major threat to human health, and practices that contribute to the development of this threat need to be addressed. Since the 1950s, antibiotics have been used in low doses to increase growth and decrease the feed requirement of animal-derived food sources. A consequence of this practice is the accelerated emergence of antimicrobial resistance that can influence human health through its distribution via animal food products. In the laboratory setting, sublethal doses of antibiotics promote the expansion of bacterial persister populations, a low energy, low metabolism phenotype characterized broadly by antibiotic tolerance. Furthermore, the induction of persister bacteria has been positively correlated with an increased emergence of antibiotic-resistant strains. This body of evidence suggests that the use of antibiotics in agriculture at subtherapeutic levels is actively catalyzing the emergence of antimicrobial-resistant bacteria through the expansion of bacterial persister populations, which is potentially leading to increased infections in humans and decreased antibiotic potency. There is an urgent need to address this debilitating effect on antibiotics and its influence on human health. In this review, we summarize the recent literature on the topic of emerging antimicrobial resistance and its association with bacterial persister populations.
Citation: Noah T Thompson, David A Kitzenberg, Daniel J Kao. Persister-mediated emergence of antimicrobial resistance in agriculture due to antibiotic growth promoters[J]. AIMS Microbiology, 2023, 9(4): 738-756. doi: 10.3934/microbiol.2023038
The creation and continued development of antibiotics have revolutionized human health and disease for the past century. The emergence of antimicrobial resistance represents a major threat to human health, and practices that contribute to the development of this threat need to be addressed. Since the 1950s, antibiotics have been used in low doses to increase growth and decrease the feed requirement of animal-derived food sources. A consequence of this practice is the accelerated emergence of antimicrobial resistance that can influence human health through its distribution via animal food products. In the laboratory setting, sublethal doses of antibiotics promote the expansion of bacterial persister populations, a low energy, low metabolism phenotype characterized broadly by antibiotic tolerance. Furthermore, the induction of persister bacteria has been positively correlated with an increased emergence of antibiotic-resistant strains. This body of evidence suggests that the use of antibiotics in agriculture at subtherapeutic levels is actively catalyzing the emergence of antimicrobial-resistant bacteria through the expansion of bacterial persister populations, which is potentially leading to increased infections in humans and decreased antibiotic potency. There is an urgent need to address this debilitating effect on antibiotics and its influence on human health. In this review, we summarize the recent literature on the topic of emerging antimicrobial resistance and its association with bacterial persister populations.
Antibiotic-Free Feed
Antibiotic Growth Promoter
Antimicrobial Peptide
Antimicrobial Resistance
Conventional Feed
Fluoroquinolone
Guidance for Industry
Minimum Duration for Killing
Minimum Inhibitory Concentration
non-Antibiotic Growth Promoter
United States Food & Drug Administration
Urinary Tract Infection

| [1] |
Hutchings MI, Truman AW, Wilkinson B (2019) Antibiotics: past, present and future. Curr Opin Microbiol 51: 72-80. https://doi.org/10.1016/j.mib.2019.10.008 
|
| [2] |
Van Boeckel TP, Pires J, Silvester R, et al. (2019) Global trends in antimicrobial resistance in animals in low- and middle-income countries. Science 365. https://doi.org/10.1126/science.aaw1944
|
| [3] | Tiseo K, Huber L, Gilbert M, et al. (2020) Global trends in antimicrobial use in food animals from 2017 to 2030. Antibiotics (Basel) 9. https://doi.org/10.3390/antibiotics9120918 |
| [4] |
Klein EY, Tseng KK, Pant S, et al. (2019) Tracking global trends in the effectiveness of antibiotic therapy using the Drug Resistance Index. BMJ Glob Health 4: e001315. https://doi.org/10.1136/bmjgh-2018-001315
|
| [5] |
Van den Bergh B, Fauvart M, Michiels J (2017) Formation, physiology, ecology, evolution and clinical importance of bacterial persisters. FEMS Microbiol Rev 41: 219-251. https://doi.org/10.1093/femsre/fux001
|
| [6] |
De Briyne N, Atkinson J, Pokludova L, et al. (2014) Antibiotics used most commonly to treat animals in Europe. Vet Rec 175. https://doi.org/10.1136/vr.102462
|
| [7] |
Manyi-Loh C, Mamphweli S, Meyer E, et al. (2018) Antibiotic use in agriculture and its consequential resistance in environmental sources: potential public health implications. Molecules 23. https://doi.org/10.3390/molecules23040795
|
| [8] |
Antimicrobial Resistance C (2022) Global burden of bacterial antimicrobial resistance in 2019: a systematic analysis. Lancet 399: 629-655. https://doi.org/10.1016/S0140-6736(21)02724-0
|
| [9] |
Blair JM, Webber MA, Baylay AJ, et al. (2015) Molecular mechanisms of antibiotic resistance. Nat Rev Microbiol 13: 42-51. https://doi.org/10.1038/nrmicro3380
|
| [10] |
Lerminiaux NA, Cameron ADS (2019) Horizontal transfer of antibiotic resistance genes in clinical environments. Can J Microbiol 65: 34-44. https://doi.org/10.1139/cjm-2018-0275
|
| [11] |
Levin-Reisman I, Ronin I, Gefen O, et al. (2017) Antibiotic tolerance facilitates the evolution of resistance. Science 355: 826-830. https://doi.org/10.1126/science.aaj2191
|
| [12] |
Levin-Reisman I, Brauner A, Ronin I, et al. (2019) Epistasis between antibiotic tolerance, persistence, and resistance mutations. Proc Natl Acad Sci USA 116: 14734-14739. https://doi.org/10.1073/pnas.1906169116
|
| [13] |
Windels EM, Michiels JE, Van den Bergh B, et al. (2019) Antibiotics: combatting tolerance to stop resistance. Mbio 10: e02095-19. https://doi.org/10.1128/mBio.02095-19
|
| [14] |
Windels EM, Michiels JE, Fauvart M, et al. (2019) Bacterial persistence promotes the evolution of antibiotic resistance by increasing survival and mutation rates. Isme J 13: 1239-1251. https://doi.org/10.1038/s41396-019-0344-9
|
| [15] |
Bigger JW (1944) The bactericidal action of penicillin on staphylococcus pyogenes. Ir J Med Sci 19: 553-568. https://doi.org/10.1007/BF02948386
|
| [16] |
Balaban NQ, Merrin J, Chait R, et al. (2004) Bacterial persistence as a phenotypic switch. Science 305: 1622-1625. https://doi.org/10.1126/science.1099390
|
| [17] |
Wilmaerts D, Windels EM, Verstraeten N, et al. (2019) General mechanisms leading to persister formation and awakening. Trends Genet 35: 401-411. https://doi.org/10.1016/j.tig.2019.03.007
|
| [18] |
Moore PR, Evenson A, et al. (1946) Use of sulfasuxidine, streptothricin, and streptomycin in nutritional studies with the chick. J Biol Chem 165: 437-441. https://doi.org/10.1016/S0021-9258(17)41154-9
|
| [19] |
Chattopadhyay MK (2014) Use of antibiotics as feed additives: a burning question. Front Microbiol 5: 334. https://doi.org/10.3389/fmicb.2014.00334
|
| [20] |
Hosain MZ, Kabir SML, Kamal MM (2021) Antimicrobial uses for livestock production in developing countries. Vet World 14: 210-221. https://doi.org/10.14202/vetworld.2021.210-221
|
| [21] |
Martin MJ, Thottathil SE, Newman TB (2015) Antibiotics overuse in animal agriculture: a call to action for health care providers. Am J Public Health 105: 2409-2410. https://doi.org/10.2105/AJPH.2015.302870
|
| [22] |
Gaskins HR, Collier CT, Anderson DB (2002) Antibiotics as growth promotants: Mode of action. Anim Biotechnol 13: 29-42. https://doi.org/10.1081/ABIO-120005768
|
| [23] |
Niewold TA (2007) The nonantibiotic anti-inflammatory effect of antimicrobial growth promoters, the real mode of action? A hypothesis. Poult Sci 86: 605-609. https://doi.org/10.1093/ps/86.4.605
|
| [24] |
Coates ME, Fuller R, Harrison GF, et al. (1963) A comparision of the growth of chicks in the Gustafsson germ-free apparatus and in a conventional environment, with and without dietary supplements of penicillin. Br J Nutr 17: 141-150. https://doi.org/10.1079/BJN19630015
|
| [25] |
Broom LJ (2017) The sub-inhibitory theory for antibiotic growth promoters. Poult Sci 96: 3104-3108. https://doi.org/10.3382/ps/pex114
|
| [26] |
Visek WJ (1978) The mode of growth promotion by antibiotics. J Anim Sci 46: 1447-1469. https://doi.org/10.2527/jas1978.4651447x
|
| [27] | Jukes TH, Williams WL (1953) Nutritional effects of antibiotics. Pharmacol Rev 5: 381-420. |
| [28] |
Kim HB, Borewicz K, White BA, et al. (2012) Microbial shifts in the swine distal gut in response to the treatment with antimicrobial growth promoter, tylosin. Proc Natl Acad Sci USA 109: 15485-15490. https://doi.org/10.1073/pnas.1205147109
|
| [29] |
Plata G, Baxter NT, Susanti D, et al. (2022) Growth promotion and antibiotic induced metabolic shifts in the chicken gut microbiome. Commun Biol 5: 293. https://doi.org/10.1038/s42003-022-03239-6
|
| [30] |
Robinson K, Becker S, Xiao Y, et al. (2019) Differential impact of subtherapeutic antibiotics and ionophores on intestinal microbiota of broilers. Microorganisms 7: 282. https://doi.org/10.3390/microorganisms7090282
|
| [31] |
Torok VA, Allison GE, Percy NJ, et al. (2011) Influence of antimicrobial feed additives on broiler commensal posthatch gut microbiota development and performance. Appl Environ Microbiol 77: 3380-3390. https://doi.org/10.1128/AEM.02300-10
|
| [32] |
Agunos A, Gow SP, Leger DF, et al. (2019) Antimicrobial use and antimicrobial resistance indicators-integration of farm-level surveillance data from broiler chickens and Turkeys in British Columbia, Canada. Front Vet Sci 6: 131. https://doi.org/10.3389/fvets.2019.00131
|
| [33] | (2017) ECDC/EFSA/EMA second joint report on the integrated analysis of the consumption of antimicrobial agents and occurrence of antimicrobial resistance in bacteria from humans and food-producing animals: Joint Interagency Antimicrobial Consumption and Resistance Analysis (JIACRA) Report. EFSA J 15: e04872. https://doi.org/10.2903/j.efsa.2017.4872 |
| [34] |
Hiki M, Kawanishi M, Abo H, et al. (2015) Decreased resistance to broad-spectrum cephalosporin in escherichia coli from healthy broilers at farms in Japan after voluntary withdrawal of ceftiofur. Foodborne Pathog Dis 12: 639-643. https://doi.org/10.1089/fpd.2015.1960
|
| [35] |
Sundsfjord A, Simonsen GS, Courvalin P (2001) Human infections caused by glycopeptide-resistant Enterococcus spp: are they a zoonosis?. Clin Microbiol Infect 7: 16-33. https://doi.org/10.1046/j.1469-0691.2001.00055.x
|
| [36] |
Aarestrup FM, Kruse H, Tast E, et al. (2000) Associations between the use of antimicrobial agents for growth promotion and the occurrence of resistance among Enterococcus faecium from broilers and pigs in Denmark, Finland, and Norway. Microb Drug Resist 6: 63-70. https://doi.org/10.1089/mdr.2000.6.63
|
| [37] |
Kumar K, Gupta SC, Chander Y, et al. (2005) Antibiotic use in agriculture and its impact on the terrestrial environment. Adv Agron 87: 1-54. https://doi.org/10.1016/S0065-2113(05)87001-4
|
| [38] |
Sarmah AK, Meyer MT, Boxall ABA (2006) A global perspective on the use, sales, exposure pathways, occurrence, fate and effects of veterinary antibiotics (VAs) in the environment. Chemosphere 65: 725-759. https://doi.org/10.1016/j.chemosphere.2006.03.026
|
| [39] |
Meyerhoff A, Albrecht R, Meyer JM, et al. (2004) US Food and Drug Administration approval of ciprofloxacin hydrochloride for management of postexposure inhalational anthrax. Clin Infect Dis 39: 303-308. https://doi.org/10.1086/421491
|
| [40] |
Chantziaras I, Boyen F, Callens B, et al. (2014) Correlation between veterinary antimicrobial use and antimicrobial resistance in food-producing animals: a report on seven countries. J Antimicrob Chem 69: 827-834. https://doi.org/10.1093/jac/dkt443
|
| [41] |
Norstrom M, Hofshagen M, Stavnes T, et al. (2006) Antimicrobial resistance in Campylobacter jejuni from humans and broilers in Norway. Epidemiol Infect 134: 127-130. https://doi.org/10.1017/S0950268805004814
|
| [42] |
de Been M, Lanza VF, de Toro M, et al. (2014) Dissemination of cephalosporin resistance genes between Escherichia coli strains from farm animals and humans by specific plasmid lineages. PLoS Genet 10: e1004776. https://doi.org/10.1371/journal.pgen.1004776
|
| [43] |
Woolhouse M, Ward M, van Bunnik B, et al. (2015) Antimicrobial resistance in humans, livestock and the wider environment. Philos Trans R Soc Lond B Biol Sci 370: 20140083. https://doi.org/10.1098/rstb.2014.0083
|
| [44] |
Muloi D, Ward MJ, Pedersen AB, et al. (2018) Are food animals responsible for transfer of antimicrobial-resistant escherichia coli or their resistance determinants to human populations? a systematic review. Foodborne Pathog Dis 15: 467-474. https://doi.org/10.1089/fpd.2017.2411
|
| [45] |
Gutierrez A, Jain S, Bhargava P, et al. (2017) Understanding and sensitizing density-dependent persistence to quinolone antibiotics. Molecular Cell 68: 1147-1154 e1143. https://doi.org/10.1016/j.molcel.2017.11.012
|
| [46] |
Helaine S, Kugelberg E (2014) Bacterial persisters: formation, eradication, and experimental systems. Trends Microbiol 22: 417-424. https://doi.org/10.1016/j.tim.2014.03.008
|
| [47] |
Stapels DAC, Hill PWS, Westermann AJ, et al. (2018) Salmonella persisters undermine host immune defenses during antibiotic treatment. Science 362: 1156-1160. https://doi.org/10.1126/science.aat7148
|
| [48] |
Balaban NQ, Helaine S, Lewis K, et al. (2019) Definitions and guidelines for research on antibiotic persistence. Nat Rev Microbiol 17: 441-448. https://doi.org/10.1038/s41579-019-0196-3
|
| [49] |
Brauner A, Fridman O, Gefen O, et al. (2016) Distinguishing between resistance, tolerance and persistence to antibiotic treatment. Nat Rev Microbiol 14: 320-330. https://doi.org/10.1038/nrmicro.2016.34
|
| [50] |
Ronneau S, Hill PW, Helaine S (2021) Antibiotic persistence and tolerance: not just one and the same. Curr Opin Microbiol 64: 76-81. https://doi.org/10.1016/j.mib.2021.09.017
|
| [51] |
Barrett TC, Mok WWK, Murawski AM, et al. (2019) Enhanced antibiotic resistance development from fluoroquinolone persisters after a single exposure to antibiotic. Nat Commun 10: 1177. https://doi.org/10.1038/s41467-019-09058-4
|
| [52] |
Allison KR, Brynildsen MP, Collins JJ (2011) Metabolite-enabled eradication of bacterial persisters by aminoglycosides. Nature 473: 216-220. https://doi.org/10.1038/nature10069
|
| [53] |
Pontes MH, Groisman EA (2019) Slow growth determines nonheritable antibiotic resistance in Salmonella enterica. Sci Signal 12. https://doi.org/10.1126/scisignal.aax3938
|
| [54] |
Fridman O, Goldberg A, Ronin I, et al. (2014) Optimization of lag time underlies antibiotic tolerance in evolved bacterial populations. Nature 513: 418-421. https://doi.org/10.1038/nature13469
|
| [55] |
Rowe SE, Wagner NJ, Li L, et al. (2020) Reactive oxygen species induce antibiotic tolerance during systemic Staphylococcus aureus infection. Nat Microbiol 5: 282-290. https://doi.org/10.1038/s41564-019-0627-y
|
| [56] | Tuomanen E, Cozens R, Tosch W, et al. (1986) The rate of killing of escherichia-coli by beta-lactam antibiotics is strictly proportional to the rate of bacterial-growth. J Gen Microbiol 132: 1297-1304. https://doi.org/10.1099/00221287-132-5-1297 |
| [57] |
Amato SM, Orman MA, Brynildsen MP (2013) Metabolic control of persister formation in Escherichia coli. Molecular Cell 50: 475-487. https://doi.org/10.1016/j.molcel.2013.04.002
|
| [58] |
Helaine S, Cheverton AM, Watson KG, et al. (2014) Internalization of Salmonella by macrophages induces formation of nonreplicating persisters. Science 343: 204-208. https://doi.org/10.1126/science.1244705
|
| [59] |
Hill PWS, Moldoveanu AL, Sargen M, et al. (2021) The vulnerable versatility of Salmonella antibiotic persisters during infection. Cell Host Microbe 29: 1757-1773. https://doi.org/10.1016/j.chom.2021.10.002
|
| [60] |
Korch SB, Henderson TA, Hill TM (2003) Characterization of the hipA7 allele of Escherichia coli and evidence that high persistence is governed by (p)ppGpp synthesis. Mol Microbiol 50: 1199-1213. https://doi.org/10.1046/j.1365-2958.2003.03779.x
|
| [61] |
Moyed HS, Bertrand KP (1983) Hipa, a newly recognized gene of escherichia-coli k-12 that affects frequency of persistence after inhibition of murein synthesis. J Bacteriol 155: 768-775. https://doi.org/10.1128/jb.155.2.768-775.1983
|
| [62] |
Wilmaerts D, Bayoumi M, Dewachter L, et al. (2018) The persistence-inducing toxin HokB forms dynamic pores that cause atp leakage. Mbio 9. https://doi.org/10.1128/mBio.00744-18
|
| [63] |
Zalis EA, Nuxoll AS, Manuse S, et al. (2019) Stochastic variation in expression of the tricarboxylic acid cycle produces persister cells. Mbio 10. https://doi.org/10.1128/mBio.01930-19
|
| [64] |
Verstraeten N, Knapen WJ, Kint CI, et al. (2015) Obg and membrane depolarization are part of a microbial bet-hedging strategy that leads to antibiotic tolerance. Mol Cell 59: 9-21. https://doi.org/10.1016/j.molcel.2015.05.011
|
| [65] |
Allison KR, Brynildsen MP, Collins JJ (2011) Heterogeneous bacterial persisters and engineering approaches to eliminate them. Curr Opin Microbiol 14: 593-598. https://doi.org/10.1016/j.mib.2011.09.002
|
| [66] |
Conlon BP (2014) Staphylococcus aureus chronic and relapsing infections: Evidence of a role for persister cells An investigation of persister cells, their formation and their role in S. aureus disease. Bioessays 36: 991-996. https://doi.org/10.1002/bies.201400080
|
| [67] |
Lewis K (2010) Persister cells. Annu Rev Microbiol 64: 357-372. https://doi.org/10.1146/annurev.micro.112408.134306
|
| [68] |
Bakkeren E, Huisman JS, Fattinger SA, et al. (2019) Salmonella persisters promote the spread of antibiotic resistance plasmids in the gut. Nature 573: 276-280. https://doi.org/10.1038/s41586-019-1521-8
|
| [69] |
Davies J, Davies D (2010) Origins and evolution of antibiotic resistance. Microbiol Mol Biol Rev 74: 417-433. https://doi.org/10.1128/MMBR.00016-10
|
| [70] |
Huddleston JR (2014) Horizontal gene transfer in the human gastrointestinal tract: potential spread of antibiotic resistance genes. Infect Drug Resist 7: 167-176. https://doi.org/10.2147/IDR.S48820
|
| [71] | Wang HQ, Feng MY, Anwar TM, et al. (2023) Change in antimicrobial susceptibility of Listeria spp. in response to stress conditions. Front Sustainable Food Systems 7. https://doi.org/10.3389/fsufs.2023.1179835 |
| [72] | Wu RA, Feng JS, Yue M, et al. (2023) Overuse of food-grade disinfectants threatens a global spread of antimicrobial-resistant bacteria. Crit Rev Food Sci Nutr . https://doi.org/10.1080/10408398.2023.2176814 |
| [73] |
Cohen NR, Lobritz MA, Collins JJ (2013) Microbial persistence and the road to drug resistance. Cell Host Microbe 13: 632-642. https://doi.org/10.1016/j.chom.2013.05.009
|
| [74] |
Douafer H, Andrieu V, Phanstiel O, et al. (2019) Antibiotic adjuvants: make antibiotics great again!. J Medl Chem 62: 8665-8681. https://doi.org/10.1021/acs.jmedchem.8b01781
|
| [75] |
Kitzenberg DA, Lee JS, Mills KB, et al. (2022) Adenosine awakens metabolism to enhance growth-independent killing of tolerant and persister bacteria across multiple classes of antibiotics. Mbio 13. https://doi.org/10.1128/mbio.00480-22
|
| [76] |
Meylan S, Andrews IW, Collins JJ (2018) Targeting antibiotic tolerance, pathogen by pathogen. Cell 172: 1228-1238. https://doi.org/10.1016/j.cell.2018.01.037
|
| [77] |
Wright GD (2016) Antibiotic adjuvants: rescuing antibiotics from resistance. Trends Microbiol 24: 862-871. https://doi.org/10.1016/j.tim.2016.06.009
|
| [78] |
Bengtsson-Palme J, Kristiansson E, Larsson DGJ (2018) Environmental factors influencing the development and spread of antibiotic resistance. Fems Microbiol Rev 42: 68-80. https://doi.org/10.1093/femsre/fux053
|
| [79] |
Blair JMA, Webber MA, Baylay AJ, et al. (2015) Molecular mechanisms of antibiotic resistance. Nat Rev Microbiol 13: 42-51. https://doi.org/10.1038/nrmicro3380
|
| [80] |
Darby EM, Trampari E, Siasat P, et al. (2023) Molecular mechanisms of antibiotic resistance revisited. Nat Rev Microbiol 21: 280-295. https://doi.org/10.1038/s41579-022-00820-y
|
| [81] |
Andersson DI, Hughes D (2014) Microbiological effects of sublethal levels of antibiotics. Nat Rev Microbiol 12: 465-478. https://doi.org/10.1038/nrmicro3270
|
| [82] |
Levy SB, Marshall B (2004) Antibacterial resistance worldwide: causes, challenges and responses. Nature Medicine 10: S122-S129. https://doi.org/10.1038/nm1145
|
| [83] | Ventola CL (2015) The antibiotic resistance crisis: part 1: causes and threats. P T 40: 277-283. |
| [84] |
Vega NM, Allison KR, Khalil AS, et al. (2012) Signaling-mediated bacterial persister formation. Nat Chem Biol 8: 431-433. https://doi.org/10.1038/nchembio.915
|
| [85] | Reeks BY, Champlin FR, Paulsen DB, et al. (2005) Effects of sub-minimum inhibitory concentration antibiotic levels and temperature on growth kinetics and outer membrane protein expression in Mannheimia haemolytica and Haemophilus somnus. Can J Vet Res 69: 1-10. |
| [86] |
Walsh SI, Peters DS, Smith PA, et al. (2019) Inhibition of protein secretion in escherichia coli and sub-mic effects of arylomycin antibiotics. Antimicrob Agents Chemother 63. https://doi.org/10.1128/AAC.01253-18
|
| [87] | MedlinePlus (n.d.) Tetracycline: Medlineplus drug information. Available from https://medlineplus.gov/druginfo/meds/a682098.html. |
| [88] | Pandey N, Cascella M Beta lactam antibiotics (2022). Available from https://www.ncbi.nlm.nih.gov/books/NBK545311/. |
| [89] | Yan A, Bryant EE Quinolones (2023). Available from https://www.ncbi.nlm.nih.gov/books/NBK557777/. |
| [90] |
Cowieson AJ, Kluenter AM (2019) Contribution of exogenous enzymes to potentiate the removal of antibiotic growth promoters in poultry production. Anim Feed Sci Technol 250: 81-92. https://doi.org/10.1016/j.anifeedsci.2018.04.026
|
| [91] |
Seal BS, Lillehoj HS, Donovan DM, et al. (2013) Alternatives to antibiotics: a symposium on the challenges and solutions for animal production. Anim Health Res Rev 14: 78-87. https://doi.org/10.1017/S1466252313000030
|
| [92] | Grassi L, Di Luca M, Maisetta G, et al. (2017) Generation of persister cells of pseudomonas aeruginosa and staphylococcus aureus by chemical treatment and evaluation of their susceptibility to membrane-targeting agents. Front Microbiol 8. https://doi.org/10.3389/fmicb.2017.01917 |
| [93] |
Hammerum AM, Heuer OE, Lester CH, et al. (2007) Comment on: withdrawal of growth-promoting antibiotics in Europe and its effects in relation to human health. Int J Antimicrob Agents 30: 466-468. https://doi.org/10.1016/j.ijantimicag.2007.07.012
|
| [94] |
Phillips I (2007) Withdrawal of growth-promoting antibiotics in Europe and its effects in relation to human health. Int J Antimicrob Agents 30: 101-107. https://doi.org/10.1016/j.ijantimicag.2007.02.018
|
| [95] |
Bywater RJ (2005) Identification and surveillance of antimicrobial resistance dissemination in animal production. Poult Sci 84: 644-648. https://doi.org/10.1093/ps/84.4.644
|
| [96] |
Casewell M, Friis C, Marco E, et al. (2003) The European ban on growth-promoting antibiotics and emerging consequences for human and animal health. J Antimicrob Chemother 52: 159-161. https://doi.org/10.1093/jac/dkg313
|
| [97] |
Acar J, Casewell M, Freeman J, et al. (2000) Avoparcin and virginiamycin as animal growth promoters: a plea for science in decision-making. Clin Microbiol Infect 6: 477-482. https://doi.org/10.1046/j.1469-0691.2000.00128.x
|
| [98] | Rahman MRT, Fliss I, Biron E (2022) Insights in the development and uses of alternatives to antibiotic growth promoters in poultry and swine production. Antibiotics (Basel) 11. https://doi.org/10.3390/antibiotics11060766 |
| [99] | U.S. Food & Drug Administration CfVMCVM GFI #213 new animal drugs and new animal drug combination products administered in or on medicated feed or drinking water of food-producing animals: recommendations for drug sponsors for voluntarily aligning product use conditions with GFI #209 (2013). Available from: https://www.fda.gov/regulatory-information/search-fda-guidance-documents/cvm-gfi-213-new-animal-drugs-and-new-animal-drug-combination-products-administered-or-medicated-feed |
| [100] | U.S. Food & Drug Administration CfVM2021 summary report on antimicrobials sold or distributed for use in food-producing animals (2022). Available from https://www.fda.gov/media/163739/download?attachment |
| [101] |
Li Y, Ed-Dra A, Tang B, et al. (2022) Higher tolerance of predominant Salmonella serovars circulating in the antibiotic-free feed farms to environmental stresses. J Hazard Mater 438: 129476. https://doi.org/10.1016/j.jhazmat.2022.129476
|

Figures(2)

Noah T Thompson, David A Kitzenberg, Daniel J Kao. Persister-mediated emergence of antimicrobial resistance in agriculture due to antibiotic growth promoters[J]. AIMS Microbiology, 2023, 9(4): 738-756. doi: 10.3934/microbiol.2023038







 DownLoad:
DownLoad: