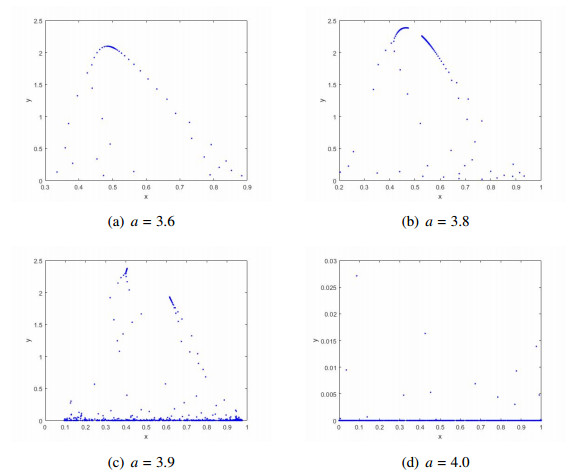
In this paper, we revisit a discrete prey-predator model with the Allee effect in prey to find its more complex dynamical properties. After pointing out and correcting those known errors for the local stability of the unique positive fixed point $ E_*, $ unlike previous studies in which the author only considered the codim 1 Neimark-Sacker bifurcation at the fixed point $ E_*, $ we focus on deriving many new bifurcation results, namely, the codim 1 transcritical bifurcation at the trivial fixed point $ E_1, $ the codim 1 transcritical and period-doubling bifurcations at the boundary fixed point $ E_2, $ the codim 1 period-doubling bifurcation and the codim 2 1:2 resonance bifurcation at the positive fixed point $ E_* $. The obtained theoretical results are also further illustrated via numerical simulations. Some new dynamics are numerically found. Our new results clearly demonstrate that the occurrence of 1:2 resonance bifurcation confirms that this system is strongly unstable, indicating that the predator and the prey will increase rapidly and breakout suddenly.
Citation: Mianjian Ruan, Xianyi Li, Bo Sun. More complex dynamics in a discrete prey-predator model with the Allee effect in prey[J]. Mathematical Biosciences and Engineering, 2023, 20(11): 19584-19616. doi: 10.3934/mbe.2023868
In this paper, we revisit a discrete prey-predator model with the Allee effect in prey to find its more complex dynamical properties. After pointing out and correcting those known errors for the local stability of the unique positive fixed point $ E_*, $ unlike previous studies in which the author only considered the codim 1 Neimark-Sacker bifurcation at the fixed point $ E_*, $ we focus on deriving many new bifurcation results, namely, the codim 1 transcritical bifurcation at the trivial fixed point $ E_1, $ the codim 1 transcritical and period-doubling bifurcations at the boundary fixed point $ E_2, $ the codim 1 period-doubling bifurcation and the codim 2 1:2 resonance bifurcation at the positive fixed point $ E_* $. The obtained theoretical results are also further illustrated via numerical simulations. Some new dynamics are numerically found. Our new results clearly demonstrate that the occurrence of 1:2 resonance bifurcation confirms that this system is strongly unstable, indicating that the predator and the prey will increase rapidly and breakout suddenly.

| [1] | A. J. Lotka, Undamped oscillations derived from the law of mass action, J. Am. Chem. Soc., 42 (1920), 1595–1599. |
| [2] |
V. Volterra, Fluctuations in the abundance of species considered Mathematically, Nature, 118 (1926), 558–560. http://dx.doi.org/10.1038/118558a0 doi: 10.1038/118558a0

|
| [3] | C. Robinson, Dynamical Systems: Stability, Symbolic dynamics, and Chaos, CRC Press, London, 1995. |
| [4] | J. M. Smith, Mathematical Ideas in Biology, Cambridge University Press, Cambridge, 1968. |
| [5] |
L. H. A. Monteiro, P. N. Mustaro, Hero's journey in bifurcation diagram, Commun, Nonl. Sci. Numer. Simul., 17 (2012), 2233–2236. https://doi.org/10.1016/j.cnsns.2011.09.035 doi: 10.1016/j.cnsns.2011.09.035
|
| [6] |
H. J. Viljoen, J. E. Gatica, H. Vladimir, Bifurcation analysis of chemically driven convection, Chem. Eng. Sci., 45 (1990), 503–517. https://doi.org/10.1016/0009-2509(90)87037-S doi: 10.1016/0009-2509(90)87037-S
|
| [7] |
C. Wang, X. Li, Further investigations into the stability and bifurcation of a discrete predator–prey model, J. Math. Anal. Appl., 422 (2015), 920–939. https://doi.org/10.1016/j.jmaa.2014.08.058 doi: 10.1016/j.jmaa.2014.08.058
|
| [8] |
S. Ruan, D. Xiao, Global analysis in a predator–prey system with nonmononotonic functional response, SIAM. J. Appl. Math., 61 (2001), 1445–1472. https://doi.org/10.1137/S0036139999361896 doi: 10.1137/S0036139999361896
|
| [9] |
O. P. Misra, P. Sinha, C. Singh, Stability and bifurcation analysis of a prey-predator model with age based predation, Appl. Math. Model., 37 (2013), 6519–6529. https://doi.org/10.1016/j.apm.2013.01.036 doi: 10.1016/j.apm.2013.01.036
|
| [10] |
S. Li, Z. Xiong, Bifurcation analysis of a predator-prey system with sex-structure and sexual favoritism, Adv. Differ. Equation, 219 (2013). https://doi.org/10.1186/1687-1847-2013-219 doi: 10.1186/1687-1847-2013-219
|
| [11] |
D. Sen, S. Ghorai, S. Sharma, M. Banerjee, Allee effect in prey's growth reduces the dynamical complexity in prey-predator model with generalist predator, Appl. Math. Model., 91 (2021), 768–790. https://doi.org/10.1016/j.apm.2020.09.046 doi: 10.1016/j.apm.2020.09.046
|
| [12] |
J. Li, G. Q. Sun, Z. Jin, Interactions of time delay and spatial diffusion induce the periodic oscillation of the vegetation system, DCDS-B, 27 (2022), 2147–2172, https://doi.org/10.3934/dcdsb.2021127 doi: 10.3934/dcdsb.2021127
|
| [13] |
P. R. Chowdhury, M. Banerjee, S. Petrovskii, Canards, relaxation oscillations, and pattern formation in a slow-fast ratio-dependent predator-prey system, Appl. Math. Model., 109 (2022), 519–535. https://doi.org/10.1016/j.apm.2022.04.022 doi: 10.1016/j.apm.2022.04.022
|
| [14] |
G. Q. Sun, L. Li, J. Li, C. Liu, Y. Wu, S. Gao, et. al, Impacts of climate change on vegetation pattern: Mathematical modeling and data analysis, Phys. Life Rev., 43 (2022), 239–270. https://doi.org/10.1016/j.plrev.2022.09.005 doi: 10.1016/j.plrev.2022.09.005
|
| [15] |
S. N. Chowdhury, J. Banerjee, M. Perc, D. Ghosh, Eco-evolutionary cyclic dominance among predators, prey, and parasites, J. Theor. Bio., 564 (2023), 111446. https://doi.org/10.1016/j.jtbi.2023.111446 doi: 10.1016/j.jtbi.2023.111446
|
| [16] |
G. Q. Sun, H. Zhang, Y. Song, L. Li, Z. Jin, Dynamic analysis of a plant-water model with spatial diffusion, JDE, 329 (2022), 395–430. https://doi.org/10.1016/j.jde.2022.05.009 doi: 10.1016/j.jde.2022.05.009
|
| [17] |
Y. Kang, A. A. Yakubu, Weak Allee effects and species coexistence, Nonl. Anal., 12 (2011), 3329–3345. https://doi.org/10.1016/j.nonrwa.2011.05.031 doi: 10.1016/j.nonrwa.2011.05.031
|
| [18] |
D. S. Boukal, L. Berec, Single-species models of the Allee effect: extinction boundaries, sex ratios and mate encounters, J. Theor. Biol., 218 (2002), 375–394. https://doi.org/10.1006/jtbi.2002.3084 doi: 10.1006/jtbi.2002.3084
|
| [19] |
C. M. Taylor, A. Hastings, Allee effects in biological invasions, Ecol. Letters, 8 (2005), 895–908. https://doi.org/10.1111/j.1461-0248.2005.00787.x doi: 10.1111/j.1461-0248.2005.00787.x
|
| [20] |
R. Sophia, J. Jang, Allee effects in a discrete-time host-parasitoid model, J. Diff. Equation Appl., 12 (2006), 165–181. https://doi.org/10.1080/10236190500539238 doi: 10.1080/10236190500539238
|
| [21] |
H. Liu, K. Zhang, Y. Ye, Y. Wei, M. Ma, Dynamic complexity and bifurcation analysis of a host–parasitoid model with Allee effect and Holling type Ⅲ functional response, Adv. Differ. Equation, 507 (2019). https://doi.org/10.1186/s13662-019-2430-8 doi: 10.1186/s13662-019-2430-8
|
| [22] |
E. Knipling, Possibilities of insect control or eradication through the use of sexually sterile males, J. Econ. Entomol, 48 (1955), 459–462. https://doi.org/10.1093/jee/48.4.459 doi: 10.1093/jee/48.4.459
|
| [23] |
B. B. Lamont, P. G. Klinkhamer, E. Witkowski, Population fragmentation may reduce fertility to zero in banksia Goodii–a demonstration of the Allee effect, Oecologia, 94 (1993), 446–450. https://doi.org/10.1007/BF00317122 doi: 10.1007/BF00317122
|
| [24] |
T. Perälä, A. Kuparinen, Detection of Allee effects in marine fishes: analytical biases generated by data availability and model selection, Proc. R. Soc. B, 284 (2017), 20171284. http://dx.doi.org/10.1098/rspb.2017.1284 doi: 10.1098/rspb.2017.1284
|
| [25] |
A. Q. Khan, Neimark–Sacker bifurcation of a two-dimensional discrete-time predator–prey model, Springer Plus 5, 126 (2016). https://doi.org/10.1186/s40064-015-1618-y doi: 10.1186/s40064-015-1618-y
|
| [26] |
F. Kangalgil, Neimark–Sacker bifurcation and stability analysis of a discrete-time prey–predator model with Allee effect in prey, Adv. Differ. Equation, 92 (2019). https://doi.org/10.1186/s13662-019-2039-y doi: 10.1186/s13662-019-2039-y
|
| [27] | S. Winggins, Introduction to Applied Nonlinear Dynamical Systems and Chaos, Springer-Verlag, New York, 2003. |
| [28] | J. Carr, Application of Center Manifold Theorem, Springer-Verlag, New York, 1981. |
| [29] |
Q. Chen, Z. Teng, F. Wang, Fold-flip and strong resonance bifurcations of a discrete-time mosquito model, Chaos Solitons Fractals, 144 (2021), 110704. https://doi.org/10.1016/j.chaos.2021.110704 doi: 10.1016/j.chaos.2021.110704
|
| [30] | Y. A. Kuznetsov, Elements of Applied Bifurcation Theory, Springer-Verlag, New York, 1998. |
| [31] |
L. G. Yuan, Q. G. Yang, Bifurcation invariant curve and hybrid control in a discrete-time predator–prey system, Appl. Math. Model, 39 (2015), 2345–2362. https://doi.org/10.1016/j.apm.2014.10.040 doi: 10.1016/j.apm.2014.10.040
|
| [32] |
B. Li, Z. M. He, Bifurcations and chaos in a two-dimensional discrete Hindmarsh–Rose model. Nonlinear Dyn., 76 (2014), 697–715. https://doi.org/10.1007/s11071-013-1161-8 doi: 10.1007/s11071-013-1161-8
|
| [33] |
B. Li, Z. M. He, Yang, 1:2 and 1:4 resonances in a two-dimensional discrete Hindmarsh-Rose model, Nonlinear Dyn., 79 (2015), 705–-720. https://doi.org/10.1007/s11071-014-1696-3 doi: 10.1007/s11071-014-1696-3
|
| [34] |
A. Singh, V. S. Sharma, Bifurcations and chaos control in a discrete-time prey–predator model with Holling type-Ⅱ functional response and prey refuge, J. Comp. Appl. Mathe., 418 (2023), 114666. https://doi.org/10.1016/j.cam.2022.114666 doi: 10.1016/j.cam.2022.114666
|
| [35] |
S. Akhtar, R. Ahmed, M. Batool, N. A. Shah, J. D. Chung, Stability, bifurcation and chaos control of a discretized Leslie prey-predator model, Chaos Solitons Fractals, 152 (2021), 111345. https://doi.org/10.1016/j.chaos.2021.111345 doi: 10.1016/j.chaos.2021.111345
|
Figures(9) / Tables(1)

Mianjian Ruan, Xianyi Li, Bo Sun. More complex dynamics in a discrete prey-predator model with the Allee effect in prey[J]. Mathematical Biosciences and Engineering, 2023, 20(11): 19584-19616. doi: 10.3934/mbe.2023868







 DownLoad:
DownLoad: