Dengue presents over 390 million cases worldwide yearly. Releasing Wolbachia-infected male mosquitoes to suppress wild mosquitoes via cytoplasmic incompatibility has proven to be a promising method for combating the disease. As cytoplasmic incompatibility causes early developmental arrest of the embryo during the larval stage, we introduce the Ricker-type survival probability to assess the resulting effects. For periodic and impulsive release strategies, our model switches between two ordinary differential equations. Owing to a Poincaré map and rigorous dynamical analyses, we give thresholds $ T^*, c^* $ and $ c^{**} (>c^*) $ for the release period $ T $ and the release amount $ c $. Then, we assume $ c > c^* $ and prove that our model admits a globally asymptotically stable periodic solution, provided $ T > T^* $, and it admits at most two periodic solutions when $ T < T^* $. Moreover, for the latter case, we assert that the origin is globally asymptotically stable if $ c\ge c^{**} $, and there exist two positive numbers such that whenever there is a periodic solution, it must initiate in an interval composed of the aforementioned two numbers, once $ c^* < c < c^{**} $. We also offer numerical examples to support the results. Finally, a brief discussion is given to evoke deeper insights into the Ricker-type model and to present our next research directions.
Citation: Zhongcai Zhu, Xue He. Rich and complex dynamics of a time-switched differential equation model for wild mosquito population suppression with Ricker-type density-dependent survival probability[J]. AIMS Mathematics, 2023, 8(12): 28670-28689. doi: 10.3934/math.20231467
Dengue presents over 390 million cases worldwide yearly. Releasing Wolbachia-infected male mosquitoes to suppress wild mosquitoes via cytoplasmic incompatibility has proven to be a promising method for combating the disease. As cytoplasmic incompatibility causes early developmental arrest of the embryo during the larval stage, we introduce the Ricker-type survival probability to assess the resulting effects. For periodic and impulsive release strategies, our model switches between two ordinary differential equations. Owing to a Poincaré map and rigorous dynamical analyses, we give thresholds $ T^*, c^* $ and $ c^{**} (>c^*) $ for the release period $ T $ and the release amount $ c $. Then, we assume $ c > c^* $ and prove that our model admits a globally asymptotically stable periodic solution, provided $ T > T^* $, and it admits at most two periodic solutions when $ T < T^* $. Moreover, for the latter case, we assert that the origin is globally asymptotically stable if $ c\ge c^{**} $, and there exist two positive numbers such that whenever there is a periodic solution, it must initiate in an interval composed of the aforementioned two numbers, once $ c^* < c < c^{**} $. We also offer numerical examples to support the results. Finally, a brief discussion is given to evoke deeper insights into the Ricker-type model and to present our next research directions.

| [1] | Dengue, World Mosquito Program, 2023. Available from: https://www.worldmosquitoprogram.org/en/learn/mosquito-borne-diseases/dengue. |
| [2] |
A. Abidemi, J. Ackora-Prah, H. O. Fatoyinbo, J. K. K. Asamoah, Lyapunov stability analysis and optimization measures for a dengue disease transmission model, Phys. A, 602 (2022), 127646. https://dx.doi.org/10.1016/j.physa.2022.127646 doi: 10.1016/j.physa.2022.127646

|
| [3] | A. Abidemi, H. O. Fatoyinbo, J. K. K. Asamoah, Analysis of dengue fever transmission dynamics with multiple controls: a mathematical approach, 2020 International Conference on Decision Aid Sciences and Application (DASA), 2020,971–978. https://dx.doi.org/10.1109/DASA51403.2020.9317064 |
| [4] |
L. Bai, L. C. Morton, Q. Liu, Climate change and mosquito-borne diseases in China: a review, Global. Health, 9 (2013), 10. https://dx.doi.org/10.1186/1744-8603-9-10 doi: 10.1186/1744-8603-9-10
|
| [5] |
L. Zou, J. Chen, X. Feng, S. Ruan, Analysis of a dengue model with vertical transmission and application to the $2014$ dengue outbreak in Guangdong province, China, Bull. Math. Biol., 80 (2018), 2633–2651. https://dx.doi.org/10.1007/s11538-018-0480-9 doi: 10.1007/s11538-018-0480-9
|
| [6] |
W. Wang, B. Yu, X. Lin, D. Kong, J. Wang, J. Tian, et al., Reemergence and autochthonous transmission of dengue virus, eastern China, $2014$, Emerg. Infec. Dis., 21 (2015), 1670–1673. https://dx.doi.org/10.3201/eid2109.150622 doi: 10.3201/eid2109.150622
|
| [7] | Dengue and severe dengue, World Health Organization, 2023. Available from: https://www.who.int/health-topics/dengue-and-severe-dengue#tab = tab_1. |
| [8] | H. F. van Emden, M. W. Service, Pest and vector management, Cambridge University Press, 2004. https://dx.doi.org/10.1017/CBO9780511616334 |
| [9] |
J. K. K. Asamoah, E. Yankson, E. Okyere, G. Sun, Z. Jin, R. Jan, et al., Optimal control and cost-effectiveness analysis for dengue fever model with asymptomatic and partial immune individuals, Results Phys., 31 (2021), 104919. https://dx.doi.org/10.1016/j.rinp.2021.104919 doi: 10.1016/j.rinp.2021.104919
|
| [10] | N. Becker, D. Petrić, M. Zgomba, C. Boase, M. Madon, C. Dahl, et al., Mosquitoes and their control, Spring-Verlag Berlin Heidelberg, 2010. https://dx.doi.org/10.1007/978-3-540-92874-4 |
| [11] |
J. Li, B. Song, X. Wang, An extended discrete Ricker population model with Allee effects, J. Differ. Equ. Appl., 13 (2007), 309–321. https://dx.doi.org/10.1080/10236190601079191 doi: 10.1080/10236190601079191
|
| [12] |
T. C. Iles, A review of stock-recruitment relationships with reference to flatfish populations, Neth. J. Sea Res., 32 (1994), 399–420. https://dx.doi.org/10.1016/0077-7579(94)90017-5 doi: 10.1016/0077-7579(94)90017-5
|
| [13] |
M. Hartmann, G. Hosack, R. Hillary, J. Vanhatalo, Gaussian process framework for temporal dependence and discrepancy functions in Ricker-type population growth models, Ann. Appl. Stat., 11 (2017), 1375–1402. https://dx.doi.org/10.1214/17-AOAS1029 doi: 10.1214/17-AOAS1029
|
| [14] |
G. Marinoschi, A. Martiradonna, Fish populations dynamics with nonlinear stock-recruitment renewal conditions, Appl. Math. Comput., 277 (2016), 101–110. https://dx.doi.org/10.1016/j.amc.2015.12.041 doi: 10.1016/j.amc.2015.12.041
|
| [15] | W. H. So, J. S. Yu, Global attractivity and uniform persistence in Nicholson's blowflies, Differ. Equ. Dyn. Syst., 2 (1994), 11–18. |
| [16] |
M. R. S. Kulenović, G. Ladas, Y. G. Sficas, Global attractivity in Nicholson's blowflies, Appl. Anal., 43 (1992), 109–124. https://dx.doi.org/10.1080/00036819208840055 doi: 10.1080/00036819208840055
|
| [17] | K. Mark, Elements of mathematical ecology, Cambridge University Press, 2001. https://dx.doi.org/10.1017/CBO9780511608520 |
| [18] | J. M. Murray, Mathematical biology, Springer-Verlag Berlin Heidelberg, 1989. https://dx.doi.org/10.1007/b98868 |
| [19] |
M. Sicard, M. Bonneau, M. Weill, Wolbachia prevalence, diversity, and ability to induce cytoplasmic incompatibility in mosquitoes, Curr. Opin. Insect Sci., 34 (2019), 12–20. https://dx.doi.org/10.1016/j.cois.2019.02.005 doi: 10.1016/j.cois.2019.02.005
|
| [20] |
R. Moretti, M. Calvitti, Issues with combining incompatible and sterile insect techniques, Nature, 590 (2021), E1–E2. https://dx.doi.org/10.1038/s41586-020-03164-w doi: 10.1038/s41586-020-03164-w
|
| [21] |
X. Zheng, D. Zhang, Y. Li, C. Yang, Y. Wu, X. Liang, et al., Incompatible and sterile insect techniques combined eliminate mosquitoes, Nature, 572 (2019), 56–61. https://dx.doi.org/10.1038/s41586-019-1407-9 doi: 10.1038/s41586-019-1407-9
|
| [22] |
T. Walker, P. Johnson, L. Moreira, I. Iturbe-Ormaetxe, F. D. Frentiu, C. J. McMeniman, et al., The wMel Wolbachia strain blocks dengue and invades caged Aedes aegypti populations, Nature, 476 (2011), 450–453. https://dx.doi.org/10.1038/nature10355 doi: 10.1038/nature10355
|
| [23] |
S. L. Dobson, W. Rattanadechakul, E. J. Marsland, Fitness advantage and cytoplasmic incompatibility in Wolbachia single- and superinfected Aedes albopictus, Heredity, 93 (2004), 135–142. https://dx.doi.org/10.1038/sj.hdy.6800458 doi: 10.1038/sj.hdy.6800458
|
| [24] |
D. Zhang, X. Zheng, Z. Xi, K. Bourtzis, J. R. L. Gilles, Combining the sterile insect technique with the incompatible insect technique: Ⅰ-impact of Wolbachia infection on the fitness of triple- and double-infected strains of Aedes albopictus, PloS One, 10 (2015), e0121126. https://dx.doi.org/10.1371/journal.pone.0121126 doi: 10.1371/journal.pone.0121126
|
| [25] |
D. Zhang, R. S. Lees, Z. Xi, K. Bourtzis, J. R. L. Gilles, Combining the sterile insect technique with the incompatible insect technique: Ⅲ-robust mating competitiveness of irradiated triple Wolbachia-infected Aedes albopictus males under semi-field conditions, PLoS One, 11 (2016), e0151864. https://dx.doi.org/10.1371/journal.pone.0151864 doi: 10.1371/journal.pone.0151864
|
| [26] |
J. Yu, Modelling mosquito population suppression based on delay differential equations, SIAM J. Appl. Math., 78 (2018), 3168–3187. https://dx.doi.org/10.1137/18M1204917 doi: 10.1137/18M1204917
|
| [27] |
J. Yu, J. Li, Global asymptotic stability in an interactive wild and sterile mosquito model, J. Differ. Equ., 269 (2020), 6193–6215. https://dx.doi.org/10.1016/j.jde.2020.04.036 doi: 10.1016/j.jde.2020.04.036
|
| [28] |
Z. Zhu, B. Zheng, Y. Shi, R. Yan, J. Yu, Stability and periodicity in a mosquito population suppression model composed of two sub-models, Nonlinear Dyn., 107 (2021), 1383–1395. https://dx.doi.org/10.1007/s11071-021-07063-1 doi: 10.1007/s11071-021-07063-1
|
| [29] |
Z. Zhu, X. Feng, L. Hu, Global dynamics of a mosquito population suppression model under a periodic release strategy, J. Appl. Anal. Comput., 13 (2023), 2297–2314. https://dx.doi.org/10.11948/20220501 doi: 10.11948/20220501
|
| [30] | J. Hale, Ordinary differential equations, New York: John Wiley, 1980. |
| [31] |
M. Han, X. Hou, L. Sheng, C. Wang, Theory of rotated equations and applications to a population model, Discrete Contin. Dyn. Syst., 38 (2018), 2171–2185. https://dx.doi.org/10.3934/dcds.2018089 doi: 10.3934/dcds.2018089
|
| [32] | Dengue-Pakistan, World Health Organization, 2022. Available from: https://www.who.int/emergencies/disease-outbreak-news/item/2022-DON414. |
| [33] | Dengue-Nepal, World Health Organization, 2022. Available from: https://www.who.int/emergencies/disease-outbreak-news/item/2022-DON412. |
| [34] |
S. B. Halstead, Dengue, Lancet, 370 (2007), 1644–1652. https://dx.doi.org/10.1016/S0140-6736(07)61687-0 doi: 10.1016/S0140-6736(07)61687-0
|
| [35] |
M. Tolle, Mosquito-borne diseases, Curr. Prob. Pediatr. Ad., 39 (2009), 97–140. https://dx.doi.org/10.1016/j.cppeds.2009.01.001 doi: 10.1016/j.cppeds.2009.01.001
|
| [36] |
N. W. Beebe, D. Pagendam, B. J. Trewin, S. A. Ritchie, Releasing incompatible males drives strong suppression across populations of wild and Wolbachia-carrying Aedes aegypti in Australia, Proc. Nati. Acad. Sci. USA, 118 (2021), e2106828118. https://dx.doi.org/10.1073/pnas.2106828118 doi: 10.1073/pnas.2106828118
|
| [37] |
J. Li, L. Cai, Y. Li, Stage-structured wild and sterile mosquito population models and their dynamics, J. Biol. Dyn., 11 (2016), 79–101. https://dx.doi.org/10.1080/17513758.2016.1159740 doi: 10.1080/17513758.2016.1159740
|
| [38] |
L. Cai, S. Ai, J. Li, Dynamics of mosquitoes populations with different strategies for releasing sterile mosquitoes, SIAM J. Appl. Math., 74 (2014), 1786–1809. https://dx.doi.org/10.1137/13094102X doi: 10.1137/13094102X
|
| [39] |
B. Zheng, J. Yu, At most two periodic solutions for a switching mosquito population suppression model, J. Dynam. Differ. Equ., 2022. https://dx.doi.org/10.1007/s10884-021-10125-y doi: 10.1007/s10884-021-10125-y
|
| [40] |
M. Huang, S. Liu, X. Song, Study of a delayed mosquito population suppression model with stage and sex structure, J. Appl. Math. Comput., 69 (2022), 89–111. https://dx.doi.org/10.1007/s12190-022-01735-w doi: 10.1007/s12190-022-01735-w
|
| [41] |
L. Hu, C. Yang, Y. Hui, J. Yu, Mosquito control based on pesticides and endosymbiotic bacterium Wolbachia, Bull. Math. Biol., 83 (2021), 58. https://dx.doi.org/10.1007/s11538-021-00881-9 doi: 10.1007/s11538-021-00881-9
|
| [42] |
Z. Zhang, B. Zheng, Dynamics of a mosquito population suppression model with a saturated Wolbachia release rate, Appl. Math. Lett., 129 (2022), 107933. https://dx.doi.org/10.1016/j.aml.2022.107933 doi: 10.1016/j.aml.2022.107933
|
| [43] |
Y. Chen, Y. Wang, J. Yu, B. Zheng, Z. Zhu, Global dynamics of a mosquito population suppression model with seasonal switching, Adv. Differ. Equ., 28 (2023), 889–920. https://dx.doi.org/10.57262/ade028-1112-889 doi: 10.57262/ade028-1112-889
|
| [44] |
J. Li, New revised simple models for interactive wild and sterile mosquito populations and their dynamics, J. Biol. Dyn., 11 (2017), 316–333. https://dx.doi.org/10.1080/17513758.2016.1216613 doi: 10.1080/17513758.2016.1216613
|
| [45] |
G. Lin, Y. Hui, Stability analysis in a mosquito population suppression model, J. Biol. Dyn., 14 (2020), 578–589. https://dx.doi.org/10.1080/17513758.2020.1792565 doi: 10.1080/17513758.2020.1792565
|
| [46] |
J. Adams, M. Jones, Estimation of lake-scale stock-recruitment models for Great Lakes sea lampreys, Ecol. Model., 467 (2022), 109916. https://dx.doi.org/10.1016/j.ecolmodel.2022.109916 doi: 10.1016/j.ecolmodel.2022.109916
|
| [47] |
Z. Liu, T. Chen, T. Zhou, Analysis of impulse release of Wolbachia to control Nilaparvata lugens, Commun. Nonlinear Sci. Numer. Simul., 116 (2023), 106842. https://dx.doi.org/10.1016/j.cnsns.2022.106842 doi: 10.1016/j.cnsns.2022.106842
|
| [48] | L. Erdős, Green heroes: from Buddha to Leonardo DiCaprio, Cham: Springer, 2019. https://dx.doi.org/10.1007/978-3-030-31806-2 |
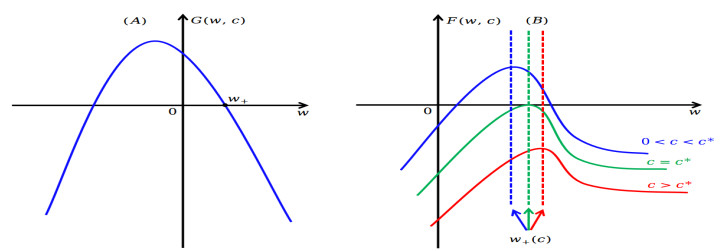
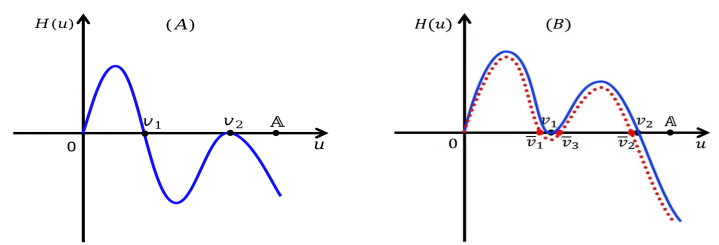
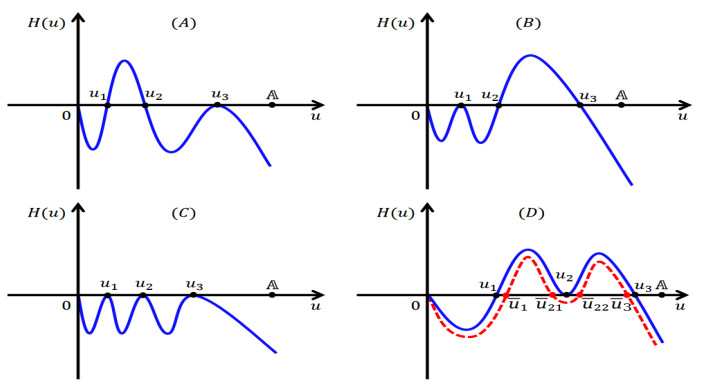
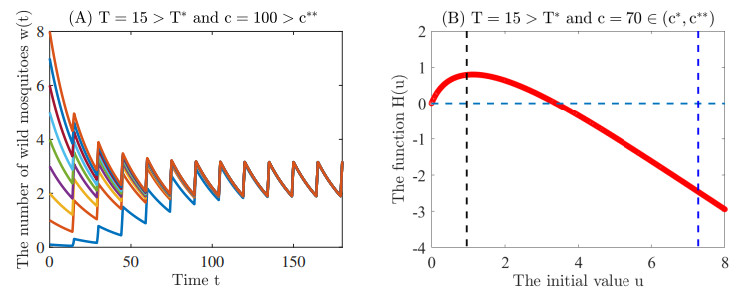
Figures(5)

Zhongcai Zhu, Xue He. Rich and complex dynamics of a time-switched differential equation model for wild mosquito population suppression with Ricker-type density-dependent survival probability[J]. AIMS Mathematics, 2023, 8(12): 28670-28689. doi: 10.3934/math.20231467







 DownLoad:
DownLoad: