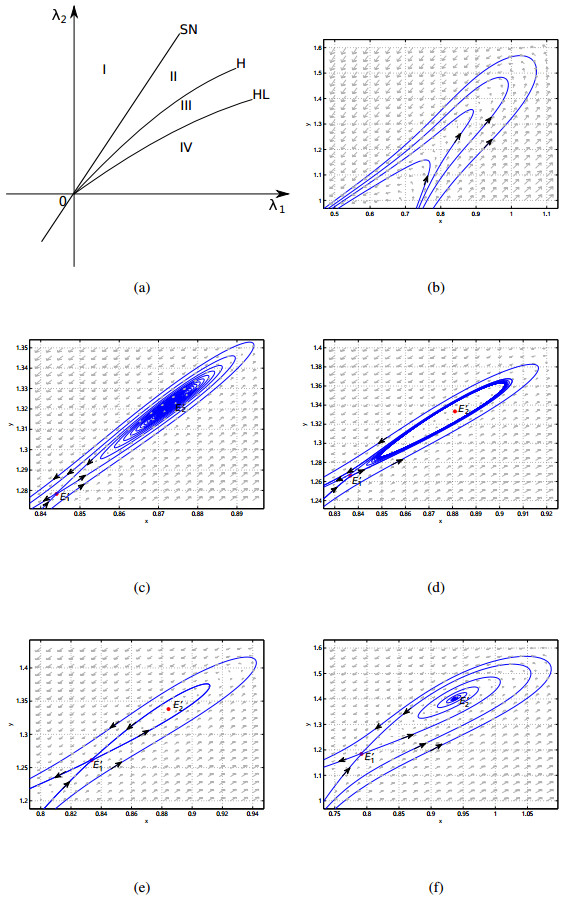
This paper is concerned with the bifurcations of a susceptible-infectious-recovered-susceptible (SIRS) epidemic model with a general saturated incidence rate $ k I^p/(1+\alpha I^p) $. For general $ p > 1 $, it is shown that the model can undergo saddle-node bifurcation, Bogdanov-Takens bifurcation of codimension two, and degenerate Hopf bifurcation of codimension two with the change of parameters. Combining with the results in [
Citation: Fang Zhang, Wenzhe Cui, Yanfei Dai, Yulin Zhao. Bifurcations of an SIRS epidemic model with a general saturated incidence rate[J]. Mathematical Biosciences and Engineering, 2022, 19(11): 10710-10730. doi: 10.3934/mbe.2022501
This paper is concerned with the bifurcations of a susceptible-infectious-recovered-susceptible (SIRS) epidemic model with a general saturated incidence rate $ k I^p/(1+\alpha I^p) $. For general $ p > 1 $, it is shown that the model can undergo saddle-node bifurcation, Bogdanov-Takens bifurcation of codimension two, and degenerate Hopf bifurcation of codimension two with the change of parameters. Combining with the results in [

| [1] |
Z. Hu, P. Bi, W. Ma, S. Ruan, Bifurcations of an SIRS epidemic model with nonlinear incidence rate, Discrete Contin. Dyn. Syst. B, 15 (2011), 93–112. https://doi.org/10.3934/dcdsb.2011.15.93 doi: 10.3934/dcdsb.2011.15.93

|
| [2] |
S. Ruan, W. Wang, Dynamical behavior of an epidemic model with a nonlinear incidence rate, J. Differ. Equations, 188 (2003), 135–163. https://doi.org/10.1016/S0022-0396(02)00089-X doi: 10.1016/S0022-0396(02)00089-X
|
| [3] |
Y. Tang, D. Huang, S. Ruan, W. Zhang, Coexistence of limit cycles and homoclinic loops in a SIRS model with a nonlinear incidence rate, SIAM J. Appl. Math., 69 (2008), 621–639. https://doi.org/10.1137/070700966 doi: 10.1137/070700966
|
| [4] |
H. W. Hethcote, The mathematics of infectious disease, SIAM Rev., 42 (2000), 599–653. https://doi.org/10.1137/S0036144500371907 doi: 10.1137/S0036144500371907
|
| [5] |
W. M. Liu, S. A. Levin, Y. Iwasa, Influence of nonlinear incidence rates upon the behavior of SIRS epidemiological models, J. Math. Biol., 23 (1986), 187–204. https://doi.org/10.1007/BF00276956 doi: 10.1007/BF00276956
|
| [6] |
V. Capasso, G. Serio, A generalization of the Kermack-McKendrick determinist epidemic model, Math. Biosci., 42 (1978), 43–61. https://doi.org/10.1016/0025-5564(78)90006-8 doi: 10.1016/0025-5564(78)90006-8
|
| [7] |
H. W. Hethcote, P. van den Driessche, Some epidemiological models with nonlinear incidence, J. Math. Biol., 29 (1991), 271–287. https://doi.org/10.1007/BF00160539 doi: 10.1007/BF00160539
|
| [8] |
W. Wang, Epidemic models with nonlinear infection forces, Math. Biosci. Eng., 3 (2006), 267–279. https://doi.org/10.3934/mbe.2006.3.267 doi: 10.3934/mbe.2006.3.267
|
| [9] |
D. Xiao, S. Ruan, Global analysis of an epidemic model with nonmonotone incidence rate, Math. Biosci., 208 (2007), 419–429. https://doi.org/10.1016/j.mbs.2006.09.025 doi: 10.1016/j.mbs.2006.09.025
|
| [10] |
G. Li, W. Wang, Bifurcation analysis of an epidemic model with nonlinear incidence, Appl. Math. Comput., 214 (2009), 411–423. https://doi.org/10.1016/j.amc.2009.04.012 doi: 10.1016/j.amc.2009.04.012
|
| [11] |
R. R. Regoes, D. Ebert, S. Bonhoeffer, Dose-dependent infection rates of parasites produce the Allee effect in epidemiology, Proc. Roy. Soc. London Ser. B, 269 (2002), 271–279. https://doi.org/10.1098/rspb.2001.1816 doi: 10.1098/rspb.2001.1816
|
| [12] |
M. G. M. Gomes, A. Margheri, G. F. Medley, C. Rebelo, Dynamical behaviour of epidemiological models with sub-optimal immunity and nonlinear incidence, J. Math. Biol., 51 (2005), 414–430. https://doi.org/10.1007/s00285-005-0331-9 doi: 10.1007/s00285-005-0331-9
|
| [13] |
M. Lu, J. Huang, S. Ruan, P. Yu, Bifurcation analysis of an SIRS epidemic model with a generalized nonmonotone and saturated incidence rate, J. Differ. Equations, 267 (2019), 1859–1898. https://doi.org/10.1016/j.jde.2019.03.005 doi: 10.1016/j.jde.2019.03.005
|
| [14] | Z. Zhang, T. Ding, W. Huang, Z. Dong, Qualitative Theory of Differential Equations, Translations of Mathematical Monographs vol. 101, American Mathematical Society, Providence, RI, 1992. |
| [15] |
J. Huang, Y. Gong, S. Ruan, Bifurcation analysis in a predator-prey model with constant-yield predator harvesting, Discrete Contin. Dyn. Syst. B, 18 (2013), 2101–2121. https://doi.org/10.3934/dcdsb.2013.18.2101 doi: 10.3934/dcdsb.2013.18.2101
|
| [16] | R. Bogdanov, Bifurcations of a limit cycle for a family of vector fields on the plane, Sel. Math. Sov., 1 (1981), 373–388. |
| [17] | R. Bogdanov, Versal deformations of a singular point on the plane in the case of zero eigen-values, Sel. Math. Sov., 1 (1981), 389–421. |
| [18] | F. Takens, Forced oscillations and bifurcation, in Applications of Global Analysis I, Communications of the Mathematical Institute Rijksuniversitat Utrecht, 3 (1974), 1–59. |
| [19] | L. Perko, Differential Equations and Dynamical System, 3rd edition, Springer, New York, 2001. |
| [20] |
Y. Dai, Y. Zhao, B. Sang, Four limit cycles in a predator-prey system of Leslie type with generalized Holling type III functional response, Nonlinear Anal. Real World Appl., 50 (2019), 218–239. https://doi.org/10.1016/j.nonrwa.2019.04.003 doi: 10.1016/j.nonrwa.2019.04.003
|
| [21] |
Y. Dai, Y. Zhao, Hopf cyclicity and global dynamics for a predator-prey system of Leslie type with simplified Holling type IV functional response, Int. J. Bifurcat. Chaos, 28 (2018), 1850166. https://doi.org/10.1142/S0218127418501663 doi: 10.1142/S0218127418501663
|


Figures(3)

Fang Zhang, Wenzhe Cui, Yanfei Dai, Yulin Zhao. Bifurcations of an SIRS epidemic model with a general saturated incidence rate[J]. Mathematical Biosciences and Engineering, 2022, 19(11): 10710-10730. doi: 10.3934/mbe.2022501







 DownLoad:
DownLoad: