Citation: Mansur Aliyu, Fatema Zohora, Ali Akbar Saboor-Yaraghi. Spleen in innate and adaptive immunity regulation[J]. AIMS Allergy and Immunology, 2021, 5(1): 1-17. doi: 10.3934/Allergy.2021001

| [1] | Greer JP, Arber DA, Glader B, et al. (2013) Wintrobes Clinical Hematology: Lymphocytes and Lymphatic Organs Philadelphia: Lippincott Williams & Wilkins, 227-410. |
| [2] |
Haley PJ (2017) The lymphoid system: a review of species differences. Toxicol Pathol 30: 111-123. doi: 10.1293/tox.2016-0075

|
| [3] |
Dale DC (2016) The mysteries of the spleen. J Leukoc Biol 100: 249-251. doi: 10.1189/jlb.4CE0216-088R
|
| [4] |
Varga I, Babala J, Kachlik DJS, et al. (2018) Anatomic variations of the spleen: current state of terminology, classification, and embryological background. Surg Radiol Anat 40: 21-29. doi: 10.1007/s00276-017-1893-0
|
| [5] |
Endo A, Ueno S, Yamada S, et al. (2015) Morphogenesis of the spleen during the human embryonic period. Anat Rec 298: 820-826. doi: 10.1002/ar.23099
|
| [6] | Ganusov VV, Tomura M (2018) Experimental and mathematical approaches to quantify recirculation kinetics of lymphocytes. BioRxiv . |
| [7] |
Mebius RE, Kraal G (2005) Structure and function of the spleen. Nat Rev Immunol 5: 606-616. doi: 10.1038/nri1669
|
| [8] | da Silva HB, Fonseca R, Pereira RM, et al. (2015) Splenic macrophage subsets and their function during blood-borne infections. Front Immunol 6: 480. |
| [9] |
Premawardena C, Bowden D, Kaplan Z, et al. (2018) Understanding of the significance and health implications of asplenia in a cohort of patients with haemaglobinopathy: possible benefits of a spleen registry. Hematology 23: 526-530. doi: 10.1080/10245332.2017.1414910
|
| [10] |
Davidson R, Wall R (2001) Prevention and management of infections in patients without a spleen. Clin Microbiol Infect 7: 657-660. doi: 10.1046/j.1198-743x.2001.00355.x
|
| [11] |
Taniguchi LU, Correia MD, Zampieri FG (2014) Overwhelming post-splenectomy infection: narrative review of the literature. Surg Infect 15: 686-693. doi: 10.1089/sur.2013.051
|
| [12] | Robier C, Schoberl M, Reiher H, et al. (2015) Bacteria on a peripheral blood smear as presenting sign of overwhelming post-splenectomy infection in a patient with secondary acute myeloid leukemia. Clin Chem Lab Med 53: e203-e205. |
| [13] |
Leone G, Pizzigallo E (2015) Bacterial infections following splenectomy for malignant and nonmalignant hematologic diseases. Mediterr J Hematol Infect Dis 7: e2015057. doi: 10.4084/mjhid.2015.057
|
| [14] |
Ugel S, Peranzoni E, Desantis G, et al. (2012) Immune tolerance to tumor antigens occurs in a specialized environment of the spleen. Cell Rep 2: 628-639. doi: 10.1016/j.celrep.2012.08.006
|
| [15] |
Salminen A, Kauppinen A, Kaarniranta K (2018) Myeloid-derived suppressor cells (MDSC): an important partner in cellular/tissue senescence. Biogerontology 19: 325-339. doi: 10.1007/s10522-018-9762-8
|
| [16] |
Hwang HK, Murakami T, Kiyuna T, et al. (2017) Splenectomy is associated with an aggressive tumor growth pattern and altered host immunity in an orthotopic syngeneic murine pancreatic cancer model. Oncotarget 8: 88827-88834. doi: 10.18632/oncotarget.21331
|
| [17] | Hwang HK, Kang CM, Lee SH, et al. (2018) Fluorescence-guided surgery with splenic preservation prevents tumor recurrence in an orthotopic nude-mouse model of human pancreatic cancer. Anticancer Res 38: 665-670. |
| [18] |
Bronte V, Pittet MJ (2013) The spleen in local and systemic regulation of immunity. Immunity 39: 806-818. doi: 10.1016/j.immuni.2013.10.010
|
| [19] |
Sty JR, Conway JJ (1985) The spleen: development and functional evaluation. Semin Nucl Med 15: 276-298. doi: 10.1016/S0001-2998(85)80005-2
|
| [20] |
Chadburn A (2000) The spleen: anatomy and anatomical function. Semin Hematol 37: 13-21. doi: 10.1016/S0037-1963(00)90113-6
|
| [21] |
Papenfuss TL, Cesta MF (2017) Spleen. Immunopathology in Toxicology and Drug Development Cham: Humana Press, 37-57. doi: 10.1007/978-3-319-47385-7_2
|
| [22] |
Kage M, Kondou R, Ogata T (2019) Anatomy of the spleen and pathology of hypersplenism. Clinical Investigation of Portal Hypertension Singapore: Springer, 25-34. doi: 10.1007/978-981-10-7425-7_3
|
| [23] |
Steiniger BS (2015) Human spleen microanatomy: why mice do not suffice. Immunology 145: 334-346. doi: 10.1111/imm.12469
|
| [24] |
den Haan JM, Kraal G (2012) Innate immune functions of macrophage subpopulations in the spleen. J Innate Immun 4: 437-445. doi: 10.1159/000335216
|
| [25] | Murphy K, Weaver C (2017) Basic concepts in immunology. Janeway's Immunobiology New York: Garland Science, 18-36. |
| [26] |
Garraud O, Borhis G, Badr G, et al. (2012) Revisiting the B-cell compartment in mouse and humans: more than one B-cell subset exists in the marginal zone and beyond. BMC Immunol 13: 63. doi: 10.1186/1471-2172-13-63
|
| [27] |
Cesta MF (2006) Normal structure, function, and histology of the spleen. Toxicol Pathol 34: 455-465. doi: 10.1080/01926230600867743
|
| [28] |
Nus M, Sage AP, Lu Y, et al. (2017) Marginal zone B cells control the response of follicular helper T cells to a high-cholesterol diet. Nat Med 23: 601-610. doi: 10.1038/nm.4315
|
| [29] |
Turner VM, Mabbott NA (2017) Ageing adversely affects the migration and function of marginal zone B cells. Immunology 151: 349-362. doi: 10.1111/imm.12737
|
| [30] |
Shlomchik MJ, Weisel F (2012) Germinal center selection and the development of memory B and plasma cells. Immunol Rev 247: 52-63. doi: 10.1111/j.1600-065X.2012.01124.x
|
| [31] |
Steiniger B, Barth P (2000) Microanatomy and Function of the Spleen Heidelberg: Springer. doi: 10.1007/978-3-642-57088-9
|
| [32] | Kumar N, Patil J, Swamy RS, et al. (2014) Atypical arterial supply to the spleen by polar branches of splenic artery and accessory splenic artery—a case report. J Clin Diagnostic Res 8: AD03-AD04. |
| [33] | Groom AC, Schmidt EE, MacDonald IC (1991) Microcirculatory pathways and blood flow in spleen: new insights from washout kinetics, corrosion casts, and quantitative intravital videomicroscopy. Scanning Microsc 5: 159-174. |
| [34] |
Steiniger BS, Ulrich C, Berthold M, et al. (2018) Capillary networks and follicular marginal zones in human spleens. Three-dimensional models based on immunostained serial sections. PLoS One 13: e0191019. doi: 10.1371/journal.pone.0191019
|
| [35] |
Pabst R (1988) The spleen in lymphocyte migration. Immunol Today 9: 43-45. doi: 10.1016/0167-5699(88)91258-3
|
| [36] | Steiniger B, Timphus EM, Jacob R, et al. (2005) CD27+ B cells in human lymphatic organs: re-evaluating the splenic marginal zone. Immunology 116: 429-442. |
| [37] |
Bickenbach KA, Gonen M, Labow DM, et al. (2013) Indications for and efficacy of splenectomy for haematological disorders. Br J Surg 100: 794-800. doi: 10.1002/bjs.9067
|
| [38] |
Coco D, Leanza S (2019) Indications for surgery in non-traumatic spleen disease. Open Access Maced J Med Sci 7: 2958-2960. doi: 10.3889/oamjms.2019.568
|
| [39] |
Fallah J, Olszewski AJ (2019) Diagnostic and therapeutic splenectomy for splenic lymphomas: analysis of the National Cancer Data Base. Hematology 24: 378-386. doi: 10.1080/16078454.2019.1591570
|
| [40] |
Huang L, Li W, Yang T, et al. (2020) Association between splenectomy and portal hypertension in the development of pulmonary hypertension. Pulm Circ 10: 1-9. doi: 10.1177/2045894019895426
|
| [41] | Nagem RG, Petroianu A (2009) Subtotal splenectomy for splenic abscess. Can J Surg 52: E91-E92. |
| [42] |
Luu S, Spelman D, Woolley IJ (2019) Post-splenectomy sepsis: preventative strategies, challenges, and solutions. Infect Drug Resist 12: 2839-2851. doi: 10.2147/IDR.S179902
|
| [43] |
Manciu S, Tudor S, Vasilescu C (2018) Splenic cysts: a strong indication for a minimally invasive partial splenectomy. Could the splenic hilar vasculature type hold a defining role? World J Surg 42: 3543-3550. doi: 10.1007/s00268-018-4650-6
|
| [44] |
Kraal G (1992) Cells in the marginal zone of the spleen. Int Rev Cytol 132: 31-74. doi: 10.1016/S0074-7696(08)62453-5
|
| [45] | Kashimura M, Shibata A (1989) Structure and functions of the human spleen: relationship between microcirculation and splenic functions. Rinsho Ketsueki 30: 1234-1238. |
| [46] |
Ram S, Lewis LA, Rice PA (2010) Infections of people with complement deficiencies and patients who have undergone splenectomy. Clin Microbiol Rev 23: 740-780. doi: 10.1128/CMR.00048-09
|
| [47] |
Sojka DK, Tian Z, Yokoyama WM (2014) Tissue-resident natural killer cells and their potential diversity. Semin Immunol 26: 127-131. doi: 10.1016/j.smim.2014.01.010
|
| [48] |
Peng H, Tian Z (2017) Diversity of tissue-resident NK cells. Semin Immunol 31: 3-10. doi: 10.1016/j.smim.2017.07.006
|
| [49] |
Spits H, Bernink JH, Lanier L (2016) NK cells and type 1 innate lymphoid cells: partners in host defense. Nat Immunol 17: 758-764. doi: 10.1038/ni.3482
|
| [50] |
Hadad U, Thauland TJ, Martinez OM, et al. (2015) NKp46 clusters at the immune synapse and regulates NK cell polarization. Front Immunol 6: 495-495. doi: 10.3389/fimmu.2015.00495
|
| [51] |
Robinette ML, Colonna M (2016) Innate lymphoid cells and the MHC. HLA 87: 5-11. doi: 10.1111/tan.12723
|
| [52] | Tomaz D, Pereira PM, Guerra N, et al. (2018) Nanoscale colocalization of NK cell activating and inhibitory receptors controls signal integration. BioRxiv . |
| [53] |
Ashouri E, Farjadian S, Reed EF, et al. (2009) KIR gene content diversity in four Iranian populations. Immunogenetics 61: 483-492. doi: 10.1007/s00251-009-0378-7
|
| [54] |
Wang HD, Jin XY, Guo YX, et al. (2019) KIR gene presence/absence polymorphisms and global diversity in the Kirgiz ethnic minority and populations distributed worldwide. Mol Biol Rep 46: 1043-1055. doi: 10.1007/s11033-018-4563-3
|
| [55] |
Osman AE, Mubasher M, ElSheikh NE, et al. (2014) Characterization of human killer immunoglobulin-like receptors (KIRs) among healthy Saudis. Hum Immunol 75: 536-540. doi: 10.1016/j.humimm.2014.02.023
|
| [56] |
Ricklin D, Reis ES, Mastellos DC, et al. (2016) Complement component C3—The “Swiss Army Knife” of innate immunity and host defense. Immunol Rev 274: 33-58. doi: 10.1111/imr.12500
|
| [57] |
Schechter MC, Satola SW, Stephens DS (2019) Host defenses to extracellular bacteria. Clin Immunol London: Elsevier, 391-402. doi: 10.1016/B978-0-7020-6896-6.00027-2
|
| [58] |
Ram S, Lewis LA, Rice PA (2010) Infections of people with complement deficiencies and patients who have undergone splenectomy. Clin Microbiol Rev 23: 740-780. doi: 10.1128/CMR.00048-09
|
| [59] |
Koppel EA, Saeland E, de Cooker DJ, et al. (2005) DC-SIGN specifically recognizes Streptococcus pneumoniae serotypes 3 and 14. Immunobiology 210: 203-210. doi: 10.1016/j.imbio.2005.05.014
|
| [60] |
Aichele P, Zinke J, Grode L, et al. (2003) Macrophages of the splenic marginal zone are essential for trapping of blood-borne particulate antigen but dispensable for induction of specific T cell responses. J Immunol 171: 1148-1155. doi: 10.4049/jimmunol.171.3.1148
|
| [61] |
Pluddemann A, Mukhopadhyay S, Sankala M, et al. (2009) SR-A, MARCO and TLRs differentially recognise selected surface proteins from Neisseria meningitidis: an example of fine specificity in microbial ligand recognition by innate immune receptors. J Innate Immun 1: 153-163. doi: 10.1159/000155227
|
| [62] | Uy PPD, Francisco DM, Trivedi A, et al. (2017) Vascular diseases of the spleen: A review. J Clin Transl Hepatol 5: 152-164. |
| [63] |
Pivkin IV, Peng Z, Karniadakis GE, et al. (2016) Biomechanics of red blood cells in human spleen and consequences for physiology and disease. Proc Natl Acad Sci USA 113: 7804-7809. doi: 10.1073/pnas.1606751113
|
| [64] |
Li H, Lu L, Li X, et al. (2018) Mechanics of diseased red blood cells in human spleen and consequences for hereditary blood disorders. Proc Natl Acad Sci USA 115: 9574-9579. doi: 10.1073/pnas.1806501115
|
| [65] |
Prasad A, Kumar SS, Dessimoz C, et al. (2013) Global regulatory architecture of human, mouse and rat tissue transcriptomes. BMC Genomics 14: 716. doi: 10.1186/1471-2164-14-716
|
| [66] |
Cook MC, Basten A, Groth BFDS (1997) Outer periarteriolar lymphoid sheath arrest and subsequent differentiation of both naive and tolerant immunoglobulin transgenic B cells is determined by B cell receptor occupancy. J Exp Med 186: 631-643. doi: 10.1084/jem.186.5.631
|
| [67] | Wluka A, Olszewski WL (2006) Innate and adaptive processes in the spleen. Ann Transplant 11: 22-29. |
| [68] |
Steinman RM, Pack M, Inaba K (1997) Dendritic cells in the T-cell areas of lymphoid organs. Immunol Rev 156: 25-37. doi: 10.1111/j.1600-065X.1997.tb00956.x
|
| [69] |
Chung JB, Silverman M, Monroe JG (2003) Transitional B cells: step by step towards immune competence. Trends Immunol 24: 343-349. doi: 10.1016/S1471-4906(03)00119-4
|
| [70] |
Chung JB, Sater RA, Fields ML, et al. (2002) CD23 defines two distinct subsets of immature B cells which differ in their responses to T cell help signals. Int Immunol 14: 157-166. doi: 10.1093/intimm/14.2.157
|
| [71] | Yilma M (2018) Review on immune tolerance mechanism and physiology. Glob J Med Res 18: 12-17. |
| [72] |
Buettner M, Bornemann M, Bode U (2013) Skin tolerance is supported by the spleen. Scand J Immunol 77: 238-245. doi: 10.1111/sji.12034
|
| [73] |
Ding J, Liu S, Zhang D, et al. (2018) Transfusion of ethylene carbodiimide-fixed donor splenocytes prolongs survival of vascularized skin allografts. J Surg Res 221: 343-352. doi: 10.1016/j.jss.2017.06.005
|
| [74] |
A-Gonzalez N, Castrillo A (2018) Origin and specialization of splenic macrophages. Cell Immunol 330: 151-158. doi: 10.1016/j.cellimm.2018.05.005
|
| [75] |
Noelia A, Guillen JA, Gallardo G, et al. (2013) The nuclear receptor LXRα controls the functional specialization of splenic macrophages. Nat Immunol 14: 831. doi: 10.1038/ni.2622
|
| [76] |
McGaha TL, Chen Y, Ravishankar B, et al. (2011) Marginal zone macrophages suppress innate and adaptive immunity to apoptotic cells in the spleen. Blood 117: 5403-5412. doi: 10.1182/blood-2010-11-320028
|
| [77] |
van Dinther D, Veninga H, Iborra S, et al. (2018) Functional CD169 on macrophages mediates interaction with dendritic cells for CD8+ T cell cross-priming. Cell Rep 22: 1484-1495. doi: 10.1016/j.celrep.2018.01.021
|
| [78] |
Grabowska J, Lopez-Venegas MA, Affandi AJ, et al. (2018) CD169(+) macrophages capture and dendritic cells instruct: The interplay of the gatekeeper and the general of the immune system. Front Immunol 9: 2472-2472. doi: 10.3389/fimmu.2018.02472
|
| [79] |
Ravishankar B, Shinde R, Liu H, et al. (2014) Marginal zone CD169+ macrophages coordinate apoptotic cell-driven cellular recruitment and tolerance. Proc Natl Acad Sci USA 111: 4215-4220. doi: 10.1073/pnas.1320924111
|
| [80] |
Miyake Y, Asano K, Kaise H, et al. (2007) Critical role of macrophages in the marginal zone in the suppression of immune responses to apoptotic cell-associated antigens. J Clin Investig 117: 2268-2278. doi: 10.1172/JCI31990
|
| [81] |
Asano K, Nabeyama A, Miyake Y, et al. (2011) CD169-positive macrophages dominate antitumor immunity by crosspresenting dead cell-associated antigens. Immunity 34: 85-95. doi: 10.1016/j.immuni.2010.12.011
|
| [82] |
Prabagar MG, Do Y, Ryu S, et al. (2012) SIGN-R1, a C-type lectin, enhances apoptotic cell clearance through the complement deposition pathway by interacting with C1q in the spleen. Cell Death Differ 20: 535-545. doi: 10.1038/cdd.2012.160
|
| [83] |
Benoit ME, Clarke EV, Morgado P, et al. (2012) Complement protein C1q directs macrophage polarization and limits inflammasome activity during the uptake of apoptotic cells. J Immunol 188: 5682-5693. doi: 10.4049/jimmunol.1103760
|
| [84] |
Morelli AE (2003) Internalization of circulating apoptotic cells by splenic marginal zone dendritic cells: dependence on complement receptors and effect on cytokine production. Blood 101: 611-620. doi: 10.1182/blood-2002-06-1769
|
| [85] |
Morelli AE, Larregina AT, Shufesky WJ, et al. (2003) Internalization of circulating apoptotic cells by splenic marginal zone dendritic cells: dependence on complement receptors and effect on cytokine production. Blood 101: 611-620. doi: 10.1182/blood-2002-06-1769
|
| [86] |
Qiu CH, Miyake Y, Kaise H, et al. (2009) Novel subset of CD8α+ dendritic cells localized in the marginal zone is responsible for tolerance to cell-associated antigens. J Immunol 182: 4127-4136. doi: 10.4049/jimmunol.0803364
|
| [87] |
Yamazaki S, Dudziak D, Heidkamp GF, et al. (2008) CD8+ CD205+ splenic dendritic cells are specialized to induce Foxp3+ regulatory T cells. J Immunol 181: 6923-6933. doi: 10.4049/jimmunol.181.10.6923
|
| [88] |
Figgett WA, Fairfax K, Vincent FB, et al. (2013) The TACI receptor regulates T-cell-independent marginal zone B cell responses through innate activation-induced cell death. Immunity 39: 573-583. doi: 10.1016/j.immuni.2013.05.019
|
| [89] |
Ren W, Grimsholm O, Bernardi AI, et al. (2015) Surrogate light chain is required for central and peripheral B-cell tolerance and inhibits anti-DNA antibody production by marginal zone B cells. Eur J Immunol 45: 1228-1237. doi: 10.1002/eji.201444917
|
| [90] |
Sonoda KH, Stein-Streilein J (2002) CD1d on antigen-transporting APC and splenic marginal zone B cells promotes NKT cell-dependent tolerance. Eur J Immunol 32: 848-857. doi: 10.1002/1521-4141(200203)32:3<848::AID-IMMU848>3.0.CO;2-I
|
| [91] |
Miles K, Simpson J, Brown S, et al. (2018) Immune tolerance to apoptotic self is mediated primarily by regulatory B1a cells. Front Immunol 8: 1962. doi: 10.3389/fimmu.2017.01952
|
| [92] |
Kohyama M, Ise W, Edelson BT, et al. (2009) Role for Spi-C in the development of red pulp macrophages and splenic iron homeostasis. Nature 457: 318-321. doi: 10.1038/nature07472
|
| [93] |
Lin HH, Faunce DE, Stacey M, et al. (2005) The macrophage F4/80 receptor is required for the induction of antigen-specific efferent regulatory T cells in peripheral tolerance. J Exp Med 201: 1615-1625. doi: 10.1084/jem.20042307
|
| [94] |
Faunce DE, Stein-Streilein J (2002) NKT cell-derived RANTES recruits APCs and CD8+ T cells to the spleen during the generation of regulatory T cells in tolerance. J Immunol 169: 31-38. doi: 10.4049/jimmunol.169.1.31
|
| [95] |
Vendomèle J, Khebizi Q, Fisson S (2017) Cellular and molecular mechanisms of anterior chamber-associated immune deviation (ACAID): What we have learned from knockout mice. Front Immunol 8: 1686-1686. doi: 10.3389/fimmu.2017.01686
|
| [96] |
D'Orazio TJ, Niederkorn JY (1998) Splenic B cells are required for tolerogenic antigen presentation in the induction of anterior chamber-associated immune deviation (ACAID). Immunology 95: 47-55. doi: 10.1046/j.1365-2567.1998.00581.x
|
| [97] |
Ashour HM, Niederkorn JY (2006) Peripheral tolerance via the anterior chamber of the eye: role of B cells in MHC class I and II antigen presentation. J Immunol 176: 5950-5957. doi: 10.4049/jimmunol.176.10.5950
|
| [98] |
Vendomèle J, Dehmani S, Khebizi Q, et al. (2018) Subretinal injection of HY peptides induces systemic antigen-specific inhibition of effector CD4(+) and CD8(+) T-cell responses. Front Immunol 9: 504. doi: 10.3389/fimmu.2018.00504
|
| [99] |
Hos D, Schaub F, Cursiefen C (2019) Does anterior chamber-associated immune deviation (ACAID) play a role in posterior lamellar keratoplasty? Case report of a splenectomized patient. BMC Ophthalmol 19: 100. doi: 10.1186/s12886-019-1114-1
|
| [100] |
Cunnusamy K, Paunicka K, Reyes N, et al. (2010) Two different regulatory T cell populations that promote corneal allograft survival. Invest Ophth Vis Sci 51: 6566-6574. doi: 10.1167/iovs.10-6161
|
| [101] |
Kunishige T, Taniguchi H, Ohno T, et al. (2019) VISTA is crucial for corneal allograft survival and maintenance of immune privilege. Invest Ophth Vis Sci 60: 4958-4965. doi: 10.1167/iovs.19-27322
|
| [102] |
Koh AS, Miller EL, Buenrostro JD, et al. (2018) Rapid chromatin repression by Aire provides precise control of immune tolerance. Nat Immunol 19: 162-172. doi: 10.1038/s41590-017-0032-8
|
| [103] |
Gardner JM, DeVoss JJ, Friedman RS, et al. (2008) Deletional tolerance mediated by extrathymic Aire-expressing cells. Science 321: 843-847. doi: 10.1126/science.1159407
|
| [104] |
Gardner James M, Metzger Todd C, McMahon Eileen J, et al. (2013) Extrathymic Aire-expressing cells are a distinct bone marrow-derived population that induce functional inactivation of CD4+ T cells. Immunity 39: 560-572. doi: 10.1016/j.immuni.2013.08.005
|
| [105] |
Ravishankar B, Liu H, Shinde R, et al. (2012) Tolerance to apoptotic cells is regulated by indoleamine 2, 3-dioxygenase. P Natl Acad Sci USA 109: 3909-3914. doi: 10.1073/pnas.1117736109
|
| [106] |
Weiberg D, Basic M, Smoczek M, et al. (2018) Participation of the spleen in the IgA immune response in the gut. PLoS One 13: e0205247. doi: 10.1371/journal.pone.0205247
|
| [107] |
Edgar LJ, Kawasaki N, Nycholat CM, et al. (2018) Targeted delivery of antigen to activated CD169+ macrophages induces bias for expansion of CD8+ T cells. Cell Chem Biol 26: 131-136. doi: 10.1016/j.chembiol.2018.10.006
|
| [108] |
Sun LM, Chen HJ, Jeng LB, et al. (2015) Splenectomy and increased subsequent cancer risk: a nationwide population-based cohort study. Am J Surg 210: 243-251. doi: 10.1016/j.amjsurg.2015.01.017
|
| [109] |
Suzuki E, Kapoor V, Jassar AS, et al. (2005) Gemcitabine selectively eliminates splenic Gr-1+/CD11b+ myeloid suppressor cells in tumor-bearing animals and enhances antitumor immune activity. Clin Cancer Res 11: 6713-6721. doi: 10.1158/1078-0432.CCR-05-0883
|
| [110] |
Imai S, Nio Y, Shiraishi T, et al. (1991) Effects of splenectomy on pulmonary metastasis and growth of SC42 carcinoma transplanted into mouse liver. J Surg Oncol 47: 178-187. doi: 10.1002/jso.2930470309
|
| [111] | Higashijima J, Shimada M, Chikakiyo M, et al. (2009) Effect of splenectomy on antitumor immune system in mice. Anticancer Res 29: 385-393. |
| [112] |
Ge YG, Gao H, Kong XT (1989) Changes of peripheral T-cell subsets in asplenic W256 tumor-bearing rats. J Surg Oncol 42: 60-68. doi: 10.1002/jso.2930420113
|
| [113] |
Davis CJ, Ilstrup DM, Pemberton JH (1988) Influence of splenectomy on survival rate of patients with colorectal cancer. Am J Surg 155: 173-179. doi: 10.1016/S0002-9610(88)80276-9
|
| [114] |
Wanebo HJ, Kennedy BJ, Winchester DP, et al. (1997) Role of splenectomy in gastric cancer surgery: adverse effect of elective splenectomy on longterm survival. J Am Coll Surg 185: 177-184. doi: 10.1016/S1072-7515(01)00901-2
|
| [115] | Fatouros M, Roukos DH, Lorenz M, et al. (2005) Impact of spleen preservation in patients with gastric cancer. Anticancer Res 25: 3023-3030. |
| [116] |
Oh SJ, Hyung WJ, Li C, et al. (2009) The effect of spleen-preserving lymphadenectomy on surgical outcomes of locally advanced proximal gastric cancer. J Surg Oncol 99: 275-280. doi: 10.1002/jso.21229
|
| [117] |
Sonoda K, Izumi K, Matsui Y, et al. (2006) Decreased growth rate of lung metastatic lesions after splenectomy in mice. Eur Surg Res 38: 469-475. doi: 10.1159/000095415
|
| [118] | Kossoy G, Ben-Hur H, Lifschitz O, et al. (2002) Mammary tumors in splenectomized rats. Oncol Rep 9: 185-188. |
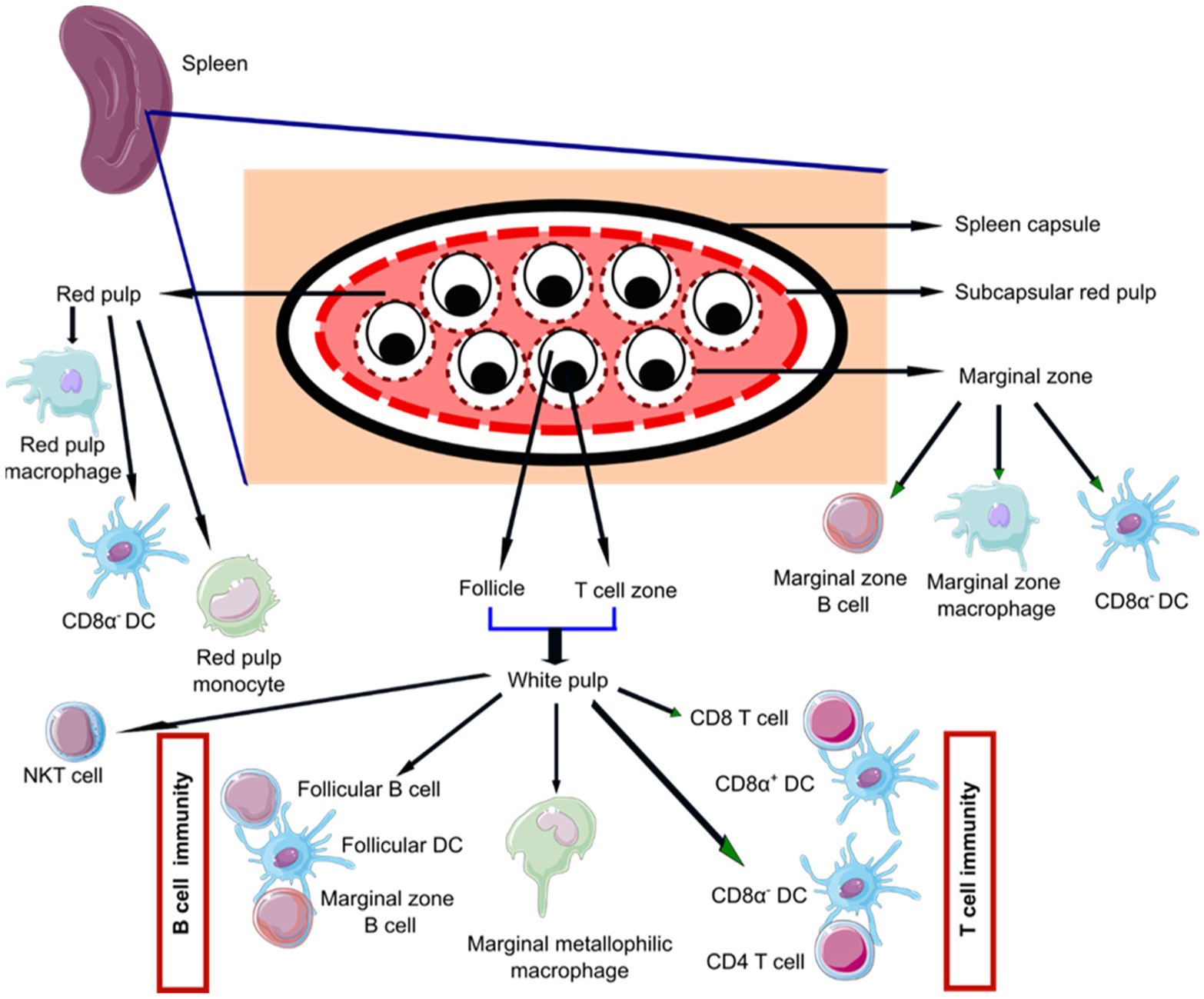

Figures(2)

Mansur Aliyu, Fatema Zohora, Ali Akbar Saboor-Yaraghi. Spleen in innate and adaptive immunity regulation[J]. AIMS Allergy and Immunology, 2021, 5(1): 1-17. doi: 10.3934/Allergy.2021001







 DownLoad:
DownLoad: