Citation: Lucia P. Pavlova, Dmitrii N. Berlov, Andres Kurismaa. Dominant and opponent relations in cortical function: An EEG study of exam performance and stress[J]. AIMS Neuroscience, 2018, 5(1): 32-55. doi: 10.3934/Neuroscience.2018.1.32

| [1] | Solomon RL, Corbit J (1974) An opponent-process theory of motivation: I. Temporal dynamics of affect. Psychol Rev 81: 119–145. |
| [2] |
Solomon RL (1980) The opponent-process theory of acquired motivation: The costs of pleasure and the benefits of pain. Am Psychol 35: 691–712. doi: 10.1037/0003-066X.35.8.691

|
| [3] | Solomon RL (1991) Acquired motivation and affective opponent-processes. In: В M. J (ed.), Neurobiology of Learning, Emotion, and Affect. New York: Raven Press Ltd., 307–347. |
| [4] | Ukhtomsky AA (1966) The Dominant, Leningrad: Leningrad State University. |
| [5] | Nadin M (Ed.) (2015) Anticipation: Learning from the Past. Cham: Springer International Publishing, 13–150. |
| [6] | Pavlova LP (2017) Dominants of the Working Brain: A Systemic Psychophysiological Approach to EEG Analysis, St.-Petersburg: Inform-Navigator. |
| [7] | AlShorman O, Ali T, Irfan M (2017) EEG Analysis for Pre-learning Stress in the Brain. In: Asian Simulation Conference. Springer, Singapore, 447–455. |
| [8] |
Robotham D, Julian C (2006) Stress and the higher education student: a critical review of the literature. J Furth High Educ 30: 107–117. doi: 10.1080/03098770600617513
|
| [9] | Strack J, Esteves F (2015) Exams? Why worry? Interpreting anxiety as facilitative and stress appraisals. Anxiety Stress Coping 28: 205–214. |
| [10] |
Lewis RS, Weekes NY, Wang TH (2007) The effect of a naturalistic stressor on frontal EEG asymmetry, stress, and health. Biol Psychol 75: 239–247. doi: 10.1016/j.biopsycho.2007.03.004
|
| [11] | Jena SK (2015) Examination stress and its effect on EEG. Int J Med Sci Pub Health 11: 1493–1497. |
| [12] |
Weekes N, Lewis R, Patel F, et al. (2006) Examination stress as an ecological inducer of cortisol and psychological responses to stress in undergraduate students. Stress 9: 199–206. doi: 10.1080/10253890601029751
|
| [13] |
Hewig, J., Schlotz, W., Gerhards, F., et al. (2008) Associations of the cortisol awakening response (CAR) with cortical activation asymmetry during the course of an exam stress period. Psychoneuroendocrinology 33: 83–91. doi: 10.1016/j.psyneuen.2007.10.004
|
| [14] |
Wiet SG, Goldstein L (1979) Successful and unsuccessful university students: Quantitative hemispheric EEG differences. Biological Psychology 8: 273–284. doi: 10.1016/0301-0511(79)90009-7
|
| [15] |
Spangler, G., Pekrun, R., Kramer, K., et al. (2002) Students' emotions, physiological reactions, and coping in academic exams. Anxiety Stress Coping 15: 413–432. doi: 10.1080/1061580021000056555
|
| [16] | Pavlova LP (2015) Individuality of brain dominants as a problem of special education and pedagogy. In: Nadin, M. (ed.) Anticipation: Learning from the Past, Cham: Springer International Publishing, 471–491. |
| [17] | Pavlova LP, Romanenko AF (1988) A Systemic Approach to Psychophysiology of the Human Brain, Leningrad: Nauka, 1988. |
| [18] | Kotchoubey B, Tretter F, Braun HA (2016) Methodological problems on the way to integrative human neuroscience. Front Integr Neurosci 10: 1–19. |
| [19] |
Krakauer, JW, Ghazanfar AA, Gomez-Marin A., et al. (2017) Neuroscience needs behavior: correcting a reductionist Bias. Neuron 93: 480–490. doi: 10.1016/j.neuron.2016.12.041
|
| [20] | Fingelkurts AA, Fingelkurts AA (2010) Short-term EEG spectral pattern as a single event in EEG phenomenology. Open Neuroimaging J 4: 130–156. |
| [21] | Fingelkurts AA, Fingelkurts AA (2015) Operational architectonics methodology for EEG analysis: theory and results. In: Sakkalis V (ed.), Modern Electroencephalographic Assessment Techniques: Theory and Applications, New York: Humana Press, 1–59. |
| [22] |
Comer CS, Harrison PK, Harrison DW (2015) The dynamic opponent relativity model: an integration and extension of capacity theory and existing theoretical perspectives on the neuropsychology of arousal and emotion. SpringerPlus 4: 345–366. doi: 10.1186/s40064-015-1120-6
|
| [23] | Luria AR (1980) Higher Cortical Functions in Man, New York: Basic Books. |
| [24] | Rusinov VS (1973) The Dominant Focus: Electrophysiological Investigations, New York: Springer Science & Business Media. |
| [25] | Kurismaa A (2015) Perspectives on Time and Anticipation in the Theory of Dominance, In: Nadin, M. (ed.), Anticipation: Learning from the Past, Cham: Springer International Publishing, 37–57. |
| [26] | Kositskiy GI, Smirnov VM (1972) The Nervous System and Stress. The principle of Dominance in Pathology (Effect of Nonspecific Stimuli on Inhibition of Pathological Processes), Washington: National Aeronautics and Space Administration; Springfield, Va. |
| [27] | Kurismaa A, Pavlova LP (2016) The Dominant as a Model of Chronogenic Change: The Relevance of AA Ukhtomsky's and LS Vygotsky's Traditions for Systemic Cognitive Studies. In Krempe SR, Smith R (eds.), Centrality of History for Theory Construction in Psychology, Germany:Springer International Publishing, 125–149. |
| [28] |
Kline JP, Blackhart GC, Williams WC (2007) Anterior EEG asymmetries and opponent process theory. Int J Psychophysiol 63: 302–307. doi: 10.1016/j.ijpsycho.2006.12.003
|
| [29] | Craig RL, Siegel PS (1979) Does negative affect beget positive affect? A test of the opponent-process theory. Bull Psycho Soc 14: 404–406. |
| [30] |
Spangler G (1997) Psychological and physiological responses during an exam and their relation to personality characteristics. Psychoneuroendocrinology 22: 423–441. doi: 10.1016/S0306-4530(97)00040-1
|
| [31] | Chanel G (2009) Emotion Assessment for Affective Computing Based on Brain and Peripheral Signals. Doctoral dissertation, Geneva: University of Geneva., 2009. |
| [32] | Ilin YP (1975) "Operational calm" and the optimum regulation of human working capacity, In Essays on the Psychology of Operator Labor, Washington, D.C: National Aeronautics and Space Administration, 226–251. |
| [33] | Jensen O, Mazaheri A (2010) Shaping functional architecture by oscillatory alpha activity: gating by inhibition. Fron Hum Neurosci 4: 1–8. |
| [34] | Klimesch W (2012) Alpha-band oscillations, attention, and controlled access to stored information, Trends Cognit Sci 16: 606–617. |
| [35] | Hanslmayr S, Staudigl T, Fellner MC (2012) Oscillatory power decreases and long-term memory: the information via desynchronization hypothesis. Front Hum Neurosci 6: 1–12. |
| [36] |
Supp GG, Siegel M, Hipp JF, et al. (2011) Cortical hypersynchrony predicts breakdown of sensory processing during loss of consciousness. Curr Biol 21: 1988–1993. doi: 10.1016/j.cub.2011.10.017
|
| [37] |
Pfurtscheller G, da Silva FHL (1999) Event-related EEG/MEG synchronization and desynchronization: basic principles. Clin Neurophysiol 110: 1842–1857. doi: 10.1016/S1388-2457(99)00141-8
|
| [38] |
Neuper C, Pfurtscheller G (2001) Event-related dynamics of cortical rhythms: frequency-specific features and functional correlates. Int J Psychophysiol 43: 41–58. doi: 10.1016/S0167-8760(01)00178-7
|
| [39] |
Neuper C, Wörtz M, Pfurtscheller G (2006) ERD/ERS patterns reflecting sensorimotor activation and deactivation. Prog Brain Res 159: 211–222. doi: 10.1016/S0079-6123(06)59014-4
|
| [40] | Sergeev GA, Pavlova LP, Romanenko AF (1968) Statistical Methods for Human Electroencephalogram Analysis, Leningrad: Nauka. |
| [41] | Doskin VA, Lavrent'eva NA, Strongina OM, et al. (1975) "SAN" psychological test applicable to studies in the field of work physiology. Gig Tr Prof Zabol 28–32. |
| [42] |
Dayan AV, Ogannisyan AO, Gevorkyan ES, et al. (2003) Reaction of cardiac activity of senior pupils of schools providing differentiated education upon examination stress. Hum Physiol 29: 160–165. doi: 10.1023/A:1022990409839
|
| [43] | Wagner EE (1983) The Hand Test: Manual, Los Angeles: Western Psychological Services. |
| [44] |
Demaree HA, Everhart DE, Youngstrom EA, et al. (2005) Brain lateralization of emotional processing: historical roots and a future incorporating "dominance". Behav Cognit Neurosci Rev 4: 3–20. doi: 10.1177/1534582305276837
|
| [45] |
Davidson RJ (2004) What does the prefrontal cortex "do" in affect: perspectives on frontal EEG asymmetry research. Biol Psychol 67: 219–234. doi: 10.1016/j.biopsycho.2004.03.008
|
| [46] |
Kelley NJ, Hortensius R, Schutter DJ, et al. (2017) The relationship of approach/avoidance motivation and asymmetric frontal cortical activity: A review of studies manipulating frontal asymmetry. Int J Psychophysiol 119: 19–30. doi: 10.1016/j.ijpsycho.2017.03.001
|
| [47] |
Groenewegen HJ, Uylings HB (2000) The prefrontal cortex and the integration of sensory, limbic and autonomic information. Prog Brain Res 126: 3–28. doi: 10.1016/S0079-6123(00)26003-2
|
| [48] |
Sarter M, Givens B, Bruno JP (2001) The cognitive neuroscience of sustained attention: where top-down meets bottom-up. Brain Res Rev 35: 146–160. doi: 10.1016/S0165-0173(01)00044-3
|
| [49] | Schulkin J (2004) Allostasis, Homeostasis, and the Costs of Physiological Adaptation, Cambridge: Cambridge University Press. |
| [50] |
Sterling P (2012) Allostasis: a model of predictive regulation. Physiol Behav 106: 5–15. doi: 10.1016/j.physbeh.2011.06.004
|
| [51] |
Schulkin J (2003) Allostasis: a neural behavioral perspective. Horm Behav 43: 21–27. doi: 10.1016/S0018-506X(02)00035-1
|
| [52] | Strack J, Lopes PN, Esteves F (2015) Will you thrive under pressure or burn out? Linking anxiety motivation and emotional exhaustion. Cognition Emotion 29: 578–591. |
| [53] | McGonigal K (2016) The Upside of Stress: Why Stress is Good for You, and How to Get Good at it. New York: Penguin. |
| [54] |
Gilbert DG, Stunkard ME, Jensen RA, et al. (1996) Effects of exam stress on mood, cortisol, and immune functioning: Influences of neuroticism and smoker-non-smoker status. Personal Individ Differences 21: 235–246. doi: 10.1016/0191-8869(96)00065-7
|
| [55] | Mega C, Ronconi L, De Beni R (2014) What makes a good student? How emotions, self-regulated learning, and motivation contribute to academic achievement. J Edu Psychol 106: 121. |
| [56] |
Pekrun R, Goetz T, Titz W, et al. (2002) Academic emotions in students' self-regulated learning and achievement: A program of qualitative and quantitative research. Edu Psychol 37: 91–105. doi: 10.1207/S15326985EP3702_4
|
| [57] | Koob GF, Le Moal M (2004) Drug addiction and allostasis. Allostasis, Homeostasis, and the Costs Physiolog Adaptation: 150–163. |
| [58] |
Ekkekakis P, Hall EE, Petruzzello SJ (2005). Variation and homogeneity in affective responses to physical activity of varying intensities: an alternative perspective on dose–response based on evolutionary considerations. J Sports Sci 23: 477–500. doi: 10.1080/02640410400021492
|
| [59] |
Woo M, Kim S, Kim J, et al. (2010) The influence of exercise intensity on frontal electroencephalographic asymmetry and self-reported affect. Res Q Exerc Sport 81: 349–359. doi: 10.1080/02701367.2010.10599683
|
| [60] |
Woo M, Kim S, Kim J, et al. (2009) Examining the exercise-affect dose–response relationship: Does duration influence frontal EEG asymmetry? Int J Psychophysiol 72: 166–172. doi: 10.1016/j.ijpsycho.2008.12.003
|
| [61] |
Regehr C, Glancy D, Pitts A (2013) Interventions to reduce stress in university students: A review and meta-analysis. J Affec Disord 148: 1–11. doi: 10.1016/j.jad.2012.11.026
|
| [62] | Flatt AK (2013) A Suffering Generation: Six Factors Contributing to the Mental Health Crisis in North American Higher Education. Coll Q 16: 1–17. |
| [63] |
Beiter R, Nash R, McCrady M, et al. (2015) The prevalence and correlates of depression, anxiety, and stress in a sample of college students. J Affect Disord 173: 90–96. doi: 10.1016/j.jad.2014.10.054
|
| [64] |
Loprinzi PD, Herod SM, Cardinal BJ (2013) Physical activity and the brain: a review of this dynamic, bi-directional relationship. Brain Res 1539: 95–104. doi: 10.1016/j.brainres.2013.10.004
|
| [65] | Heijnen S, Hommel B, Kibele A, et al. (2016) Neuromodulation of aerobic exercise-a review. Front Psychol 6: 1890-1896. |
| [66] |
Hanslmayr S, Sauseng P, Doppelmayr M (2005) Increasing individual upper alpha power by neurofeedback improves cognitive performance in human subjects. Appl Psychophysiol Biofeedback 30: 1–10. doi: 10.1007/s10484-005-2169-8
|
| [67] |
Gruzelier JH (2014) EEG-neurofeedback for optimising performance: A review of cognitive and affective outcome in healthy participants. Neurosci Biobehav Revi 44: 124–141. doi: 10.1016/j.neubiorev.2013.09.015
|
| [68] |
Ratanasiripong P, Sverduk K, Prince J, et al. (2012) Biofeedback and counseling for stress and anxiety among college students. J Coll Stud Dev 53: 742–749. doi: 10.1353/csd.2012.0070
|
| [69] | Astafurov VI, Pavlova LP (1981) Some features of alpha-activity self-control through visual feedback. All-Russian Inst Sci Tech Inf: 4214–4281. |
| [70] |
Molenaar PC (2007) Psychological methodology will change profoundly due to the necessity to focus on intra-individual variation. Integrative Psychol Behav Sci 41: 35–40. doi: 10.1007/s12124-007-9011-1
|
| [71] | de Jong T, Van Gog T, Jenks K, et al. (2009) Explorations in Learning and the Brain: On the Potential of Cognitive Neuroscience for Educational Science. New York: Springer Science & Business Media. |
| [72] |
Stein Z, Fischer KW (2011) Directions for mind, brain, and education: Methods, models, and morality. Edu Philos Theory 43: 56–66. doi: 10.1111/j.1469-5812.2010.00708.x
|
| [73] |
de-Wit L, Alexander D, Ekroll V, et al. (2016) Is neuroimaging measuring information in the brain? Psychon Bull Rev 23: 1415–1429. doi: 10.3758/s13423-016-1002-0
|
| [74] |
Barry RJ, Clarke AR, Johnstone SJ (2007) EEG differences between eyes-closed and eyes-open resting conditions. Clin Neurophysiol 118: 2765–2773. doi: 10.1016/j.clinph.2007.07.028
|
| [75] |
Whitham EM, Pope KJ, Fitzgibbon SP (2007) Scalp electrical recording during paralysis: quantitative evidence that EEG frequencies above 20Hz are contaminated by EMG. Clin Neurophysiol 118: 1877–1888. doi: 10.1016/j.clinph.2007.04.027
|
| [76] | Corning HK (1946) Lehrbuch der Topographischen Anatomie. Berlin Heidelberg: Springer. |
| [77] | Mattson MP, Calabrese EJ (2014) Hormesis. A Revolution in Biology, Toxicology and Medicine. The United States: Humana Press. |
| [78] | Nasonov DN (1962) Local Reaction of Protoplasm and Gradual Excitation. Washington D.C: National Science Foundation. |
| [79] | Matveev VV (2005) Protoreaction of protoplasm. Cell Mol Biol 51: 715–723. |
| [80] |
Agutter PS (2007) Cell mechanics and stress: from molecular details to the 'universal cell reaction' and hormesis. BioEssays 29: 324–333. doi: 10.1002/bies.20550
|
| [81] |
Agutter PS (2008) Elucidating the mechanism (s) of hormesis at the cellular level: the universal cell response. Am J Pharmacol Toxicol 3: 100–110. doi: 10.3844/ajptsp.2008.100.110
|
| [82] |
Mattson MP (2008) Awareness of hormesis will enhance future research in basic and applied neuroscience. Crit Rev Toxicol 38: 633–639. doi: 10.1080/10408440802026406
|
| [83] |
Calabrese EJ (2008) Neuroscience and hormesis: overview and general findings. Crit Rev Toxicol 38: 249–252. doi: 10.1080/10408440801981957
|
| [84] |
Daw ND, Kakade S, Dayan P (2002) Opponent interactions between serotonin and dopamine. Neural Networks 15: 603–616. doi: 10.1016/S0893-6080(02)00052-7
|
| [85] |
Boureau YL, Dayan P (2011) Opponency revisited: competition and cooperation between dopamine and serotonin. Neuropsychopharmacology 36: 74–97. doi: 10.1038/npp.2010.151
|
| [86] | Tsitolovsky LE (2015) Endogenous Generation of Goals and Homeostasis. In: Anticipation: Learning from the Past , Cham: Springer International Publishing, 175–191. |
| [87] |
Viru A (2002) Early contributions of Russian stress and exercise physiologists. J Appl Physiol 92: 1378–1382. doi: 10.1152/japplphysiol.00435.2001
|
| [88] |
Pavlova LP (2015) Work capacity and anticipation in AA Ukhtomsky's concept of dominance. Int J General Syst 44: 667–685. doi: 10.1080/03081079.2015.1032528
|
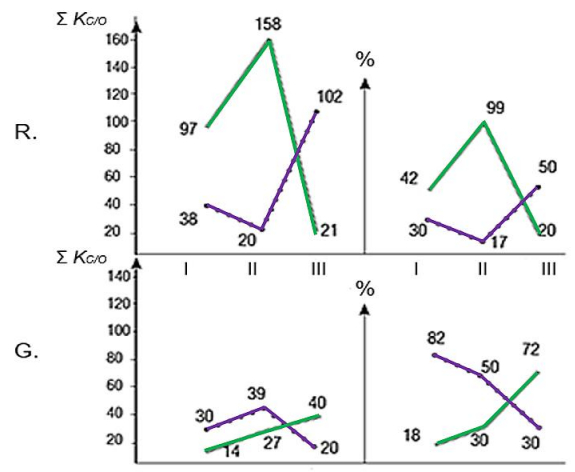
Figures(5) / Tables(2)

Lucia P. Pavlova, Dmitrii N. Berlov, Andres Kurismaa. Dominant and opponent relations in cortical function: An EEG study of exam performance and stress[J]. AIMS Neuroscience, 2018, 5(1): 32-55. doi: 10.3934/Neuroscience.2018.1.32







 DownLoad:
DownLoad: