Citation: Yue Tang, Shaun Cawthraw, Mary C. Bagnall, Adriana J. Gielbert, Martin J. Woodward, Liljana Petrovska. Identification of temperature regulated factors of Campylobacter jejuni and their potential roles in virulence[J]. AIMS Microbiology, 2017, 3(4): 885-898. doi: 10.3934/microbiol.2017.4.885

| [1] | Silva J, Leite D, Fernandes M, et al. (2011) Campylobacter spp. as a foodborne pathogen: A review. Front Microbiol 2: 200. |
| [2] | van Vliet AH, Ketley JM (2001) Pathogenesis of enteric Campylobacter infection. J Appl Microbiol: 45S–56S. |
| [3] |
Backert S, Boehm M, Wessler S, et al. (2013) Transmigration route of Campylobacter jejuni across polarized intestinal epithelial cells: paracellular, transcellular or both? Cell Commun Signal 11: 72. doi: 10.1186/1478-811X-11-72

|
| [4] |
Hermans D, Deun KV, Martel A, et al. (2011) Colonization factors of Campylobacter jejuni in the chicken gut. Vet Res 42: 82. doi: 10.1186/1297-9716-42-82
|
| [5] |
Smith CK, Abuoun M, Cawthraw SA, et al. (2008) Campylobacter colonization of the chicken induces a proinflammatory response in mucosal tissues. FEMS Immunol Med Mic 54: 114–121. doi: 10.1111/j.1574-695X.2008.00458.x
|
| [6] | Humphrey S, Chaloner G, Kemmett K, et al. (2014) Campylobacter jejuni is not merely a commensal in commercial broiler chickens and affects bird welfare. Mbio 5: e01364. |
| [7] |
Berrang ME, Buhr RJ, Cason JA, et al. (2001) Broiler carcass contamination with Campylobacter from feces during defeathering. J Food Protect 64: 2063–2066. doi: 10.4315/0362-028X-64.12.2063
|
| [8] |
European Food Safety A (2010) Scientific opinion on quantification of the risk posed by broiler meat to human campylobacteriosis in the EU. EFSA J 8: 1437. doi: 10.2903/j.efsa.2010.1437
|
| [9] |
Skirrow MB, Benjamin J (1980) '1001' Campylobacters: cultural characteristics of intestinal campylobacters from man and animals. J Hyg 85: 427–442. doi: 10.1017/S0022172400063506
|
| [10] |
Cawthraw SA, Wassenaar TM, Ayling R, et al. (1996) Increased colonization potential of Campylobacter jejuni strain 81116 after passage through chickens and its implication on the rate of transmission within flocks. Epidemiol Infect 117: 213–215. doi: 10.1017/S0950268800001333
|
| [11] |
van Doorn PA, Ruts L, Jacobs BC (2008) Clinical features, pathogenesis, and treatment of Guillain-Barre syndrome. Lancet Neurol 7: 939–950. doi: 10.1016/S1474-4422(08)70215-1
|
| [12] |
Thies FL, Hartung HP, Giegerich G (1998) Cloning and expression of the Campylobacter jejuni lon gene detected by RNA arbitrarily primed PCR. FEMS Microbiol Lett 165: 329–334. doi: 10.1111/j.1574-6968.1998.tb13165.x
|
| [13] | Thies FL, Karch H, Hartung HP, et al. (1999) Cloning and expression of the dnaK gene of Campylobacter jejuni and antigenicity of heat shock protein 70. Infect Immun 67: 1194–1200. |
| [14] |
Thies FL, Karch H, Hartung HP, et al. (1999) The ClpB protein from Campylobacter jejuni: molecular characterization of the encoding gene and antigenicity of the recombinant protein. Gene 230: 61–67. doi: 10.1016/S0378-1119(99)00054-2
|
| [15] |
Thies FL, Weishaupt A, Karch H, et al. (1999) Cloning, sequencing and molecular analysis of the Campylobacter jejuni groESL bicistronic operon. Microbiology 145: 89–98. doi: 10.1099/13500872-145-1-89
|
| [16] | Konkel ME, Kim BJ, Klena JD, et al. (1998) Characterization of the thermal stress response of Campylobacter jejuni. Infect Immun 66: 3666–3672. |
| [17] |
Bukau B (1993) Regulation of the Escherichia coli heat-shock response. Mol Microbiol 9: 671–680. doi: 10.1111/j.1365-2958.1993.tb01727.x
|
| [18] |
Skirrow MB (1977) Campylobacter enteritis: a "new" disease. Brit Med J 2: 9–11. doi: 10.1136/bmj.2.6078.9
|
| [19] |
Senior NJ, Bagnall MC, Champion OL, et al. (2011) Galleria mellonella as an infection model for Campylobacter jejuni virulence. J Med Microbiol 60: 661–669. doi: 10.1099/jmm.0.026658-0
|
| [20] |
Parkhill J, Wren BW, Mungall K, et al. (2000) The genome sequence of the food-borne pathogen Campylobacter jejuni reveals hypervariable sequences. Nature 403: 665–668. doi: 10.1038/35001088
|
| [21] |
Gaynor EC, Cawthraw S, Manning G, et al. (2004) The genome-sequenced variant of Campylobacter jejuni NCTC 11168 and the original clonal clinical isolate differ markedly in colonization, gene expression, and virulence-associated phenotypes. J Bacteriol 186: 503–517. doi: 10.1128/JB.186.2.503-517.2004
|
| [22] |
Tang Y, Underwood A, Gielbert A, et al. (2014) Metaproteomics analysis reveals the adaptation process for the chicken gut microbiota. Appl Environ Microb 80: 478–485. doi: 10.1128/AEM.02472-13
|
| [23] |
Wisniewski JR, Zougman A, Mann M (2009) Combination of FASP and StageTip-based fractionation allows in-depth analysis of the hippocampal membrane proteome. J Proteome Res 8: 5674–5678. doi: 10.1021/pr900748n
|
| [24] |
Polpitiya AD, Qian WJ, Jaitly N, et al. (2008) DAnTE: a statistical tool for quantitative analysis of -omics data. Bioinformatics 24: 1556–1558. doi: 10.1093/bioinformatics/btn217
|
| [25] | Champion OL, Karlyshev AV, Senior NJ, et al. (2010) Insect infection model for Campylobacter jejuni reveals that O-methyl phosphoramidate has insecticidal activity. J Infect Dis 201: 776–782. |
| [26] |
Atashpaz-Gargari E, Braga-Neto UM, Dougherty ER (2014) Modeling and systematic analysis of biomarker validation using selected reaction monitoring. Eurasip J Bioinform Syst Biol 2014: 17. doi: 10.1186/s13637-014-0017-y
|
| [27] |
Mermelekas G, Vlahou A, Zoidakis J (2015) SRM/MRM targeted proteomics as a tool for biomarker validation and absolute quantification in human urine. Expert Rev Mol Diagn 15: 1441–1454. doi: 10.1586/14737159.2015.1093937
|
| [28] |
Pajaniappan M, Hall JE, Cawthraw SA, et al. (2008) A temperature-regulated Campylobacter jejuni gluconate dehydrogenase is involved in respiration-dependent energy conservation and chicken colonization. Mol Microbiol 68: 474–491. doi: 10.1111/j.1365-2958.2008.06161.x
|
| [29] |
Stintzi A (2003) Gene expression profile of Campylobacter jejuni in response to growth temperature variation. J Bacteriol 185: 2009–2016. doi: 10.1128/JB.185.6.2009-2016.2003
|
| [30] |
Bahl H, Muller H, Behrens S, et al. (1995) Expression of heat shock genes in Clostridium acetobutylicum. FEMS Microbiol Rev 17: 341–348. doi: 10.1111/j.1574-6976.1995.tb00217.x
|
| [31] |
Arsene F, Tomoyasu T, Bukau B (2000) The heat shock response of Escherichia coli. Int J Food Microbiol 55: 3–9. doi: 10.1016/S0168-1605(00)00206-3
|
| [32] |
De AM, Di CR, Huet C, et al. (2004) Heat shock response in Lactobacillus plantarum. Appl Environ Microb 70: 1336–1346. doi: 10.1128/AEM.70.3.1336-1346.2004
|
| [33] | Zhang MJ, Xiao D, Zhao F, et al. (2009) Comparative proteomic analysis of Campylobacter jejuni cultured at 37 °C and 42 °C. Jpn J Infect Dis 62: 356–361. |
| [34] |
Fonseca BB, Rossi DA, Maia CA, et al. (2014) Characterization of the virulence, growth temperature and antibiotic resistance of the Campylobacter jejuni IAL 2383 strain isolated from humans. Braz J Microbiol 45: 271–274. doi: 10.1590/S1517-83822014000100039
|
| [35] | Babakhani FK, Joens LA (1993) Primary swine intestinal cells as a model for studying Campylobacter jejuni invasiveness. Infect Immun 61: 2723–2726. |
| [36] | Fox JG, Ackerman JI, Taylor N, et al. (1987) Campylobacter jejuni infection in the ferret: an animal model of human campylobacteriosis. Am J Vet Res 48: 85–90. |
| [37] | Newell DG (2001) Animal models of Campylobacter jejuni colonization and disease and the lessons to be learned from similar Helicobacter pylori models. Symp Ser Soc Appl Microbiol: 57S–67S. |
| [38] |
McLaughlin SD, Walker AW, Churcher C, et al. (2010) The bacteriology of pouchitis: a molecular phylogenetic analysis using 16S rRNA gene cloning and sequencing. Ann Surg 252: 90–98. doi: 10.1097/SLA.0b013e3181e3dc8b
|
| [39] |
Viegas SC, Mil-Homens D, Fialho AM, et al. (2013) The virulence of Salmonella enterica Serovar Typhimurium in the insect model galleria mellonella is impaired by mutations in RNase E and RNase III. Appl Environ Microb 79: 6124–6133. doi: 10.1128/AEM.02044-13
|
| [40] |
Chadwick JS, Caldwell SS, Chadwick P (1990) Adherence patterns and virulence for Galleria mellonella larvae of isolates of Serratia marcescens. J Invertebr Pathol 55: 133–134. doi: 10.1016/0022-2011(90)90044-7
|
| [41] |
Purves J, Cockayne A, Moody PC, et al. (2010) Comparison of the regulation, metabolic functions, and roles in virulence of the glyceraldehyde-3-phosphate dehydrogenase homologues gapA and gapB in Staphylococcus aureus. Infect Immun 78: 5223–5232. doi: 10.1128/IAI.00762-10
|
| [42] |
Jander G, Rahme LG, Ausubel FM (2000) Positive correlation between virulence of Pseudomonas aeruginosa mutants in mice and insects. J Bacteriol 182: 3843–3845. doi: 10.1128/JB.182.13.3843-3845.2000
|
| [43] |
Mylonakis E, Moreno R, El Khoury JB, et al. (2005) Galleria mellonella as a model system to study Cryptococcus neoformans pathogenesis. Infect Immun 73: 3842–3850. doi: 10.1128/IAI.73.7.3842-3850.2005
|
| [44] |
Kimbrell DA, Beutler B (2001) The evolution and genetics of innate immunity. Nat Rev Genet 2: 256–267. doi: 10.1038/35066006
|
| [45] |
Kavanagh K, Reeves EP (2004) Exploiting the potential of insects for in vivo pathogenicity testing of microbial pathogens. FEMS Microbiol Rev 28: 101–112. doi: 10.1016/j.femsre.2003.09.002
|
| [46] |
Gundogdu O, Mills DC, Elmi A, et al. (2011) The Campylobacter jejuni transcriptional regulator Cj1556 plays a role in the oxidative and aerobic stress response and is important for bacterial survival in vivo. J Bacteriol 193: 4238–4249. doi: 10.1128/JB.05189-11
|
| [47] |
Humphrey S, Lacharme-Lora L, Chaloner G, et al. (2015) Heterogeneity in the infection biology of Campylobacter jejuni isolates in three infection models reveals an invasive and virulent phenotype in a ST21 isolate from poultry. PLoS One 10: e0141182. doi: 10.1371/journal.pone.0141182
|
| [48] | Tsakas S, Marmaras VJ (2010) Insect immunity and its signalling: an overview. ISJ-Invert Surviv J 7: 228–238. |
| [49] |
Wallin RP, Lundqvist A, More SH, et al. (2002) Heat-shock proteins as activators of the innate immune system. Trends Immunol 23: 130–135. doi: 10.1016/S1471-4906(01)02168-8
|
| [50] |
Fujimoto S, Fujita M (1999) Serum antibody level against GroEL type heat-shock protein of Campylobacter jejuni in patients with Guillain-Barre syndrome. Microbiol Immunol 43: 241–244. doi: 10.1111/j.1348-0421.1999.tb02399.x
|
| [51] |
Akira S, Uematsu S, Takeuchi O (2006) Pathogen recognition and innate immunity. Cell 124: 783–801. doi: 10.1016/j.cell.2006.02.015
|
| [52] |
Bolla JM, De E, Dorez A, et al. (2000) Purification, characterization and sequence analysis of Omp50,a new porin isolated from Campylobacter jejuni. Biochem J 352: 637–643. doi: 10.1042/bj3520637
|
| [53] |
Dedieu L, Pages JM, Bolla JM (2008) The omp50 gene is transcriptionally controlled by a temperature-dependent mechanism conserved among thermophilic Campylobacter species. Res Microbiol 159: 270–278. doi: 10.1016/j.resmic.2008.03.002
|
| [54] |
Stancik LM, Stancik DM, Schmidt B, et al. (2002) pH-dependent expression of periplasmic proteins and amino acid catabolism in Escherichia coli. J Bacteriol 184: 4246–4258. doi: 10.1128/JB.184.15.4246-4258.2002
|
| [55] |
Handa N, Terada T, Doi-Katayama Y, et al. (2005) Crystal structure of a novel polyisoprenoid-binding protein from Thermus thermophilus HB8. Protein Sci 14: 1004–1010. doi: 10.1110/ps.041183305
|
| [56] | Patskovsky Y, Ramagopal U, Almo SC (2005) Crystal structure of campylobacter jejuni ycei periplasmic protein. NCBI Molecular Modeling Database (MMDB). |
| [57] |
Sisinni L, Cendron L, Favaro G, et al. (2010) Helicobacter pylori acidic stress response factor HP1286 is a YceI homolog with new binding specificity. FEBS J 277: 1896–1905. doi: 10.1111/j.1742-4658.2010.07612.x
|
| [58] |
Weber A, Kogl SA, Jung K (2006) Time-dependent proteome alterations under osmotic stress during aerobic and anaerobic growth in Escherichia coli. J Bacteriol 188: 7165–7175. doi: 10.1128/JB.00508-06
|
| [59] |
El-Halfawy OM, Valvano MA (2013) Chemical communication of antibiotic resistance by a highly resistant subpopulation of bacterial cells. PLoS One 8: e68874. doi: 10.1371/journal.pone.0068874
|
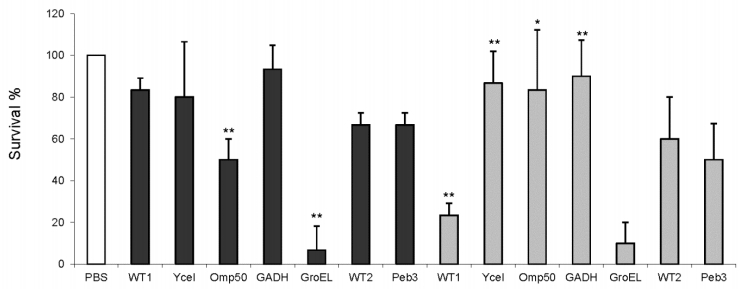
Figures(1) / Tables(2)

Yue Tang, Shaun Cawthraw, Mary C. Bagnall, Adriana J. Gielbert, Martin J. Woodward, Liljana Petrovska. Identification of temperature regulated factors of Campylobacter jejuni and their potential roles in virulence[J]. AIMS Microbiology, 2017, 3(4): 885-898. doi: 10.3934/microbiol.2017.4.885







 DownLoad:
DownLoad: