Citation: Samantha B. Gacasan, Daniel L. Baker, Abby L. Parrill. G protein-coupled receptors: the evolution of structural insight[J]. AIMS Biophysics, 2017, 4(3): 491-527. doi: 10.3934/biophy.2017.3.491

| [1] |
Venter JC, Adams MD, Myers EW, et al. (2001) The sequence of the human genome. Science 291: 1304–1351. doi: 10.1126/science.1058040

|
| [2] |
Lander ES, Linton LM, Birren B, et al. (2001) Initial sequencing and analysis of the human genome. Nature 409: 860–921. doi: 10.1038/35057062
|
| [3] | Fredriksson R, Lagerstrom MC, Lundin LG, et al. (2003) The G-protein-coupled receptors in the human genome form five main families. Phylogenetic analysis, paralogon groups, and fingerprints. Mol Pharmacol 63: 1256–1272. |
| [4] |
Pierce KL, Premont RT, Lefkowitz RJ (2002) Seven-transmembrane receptors. Nat Rev Mol Cell Biol 3: 639–650. doi: 10.1038/nrm908
|
| [5] |
Lagerstrom MC, Schioth HB (2008) Structural diversity of G protein-coupled receptors and significance for drug discovery. Nat Rev Drug Discov 7: 339–357. doi: 10.1038/nrd2518
|
| [6] |
Chien EY, Liu W, Zhao Q, et al. (2010) Structure of the human dopamine D3 receptor in complex with a D2/D3 selective antagonist. Science 330: 1091–1095. doi: 10.1126/science.1197410
|
| [7] |
Yang J, Villar VA, Armando I, et al. (2016) G protein-coupled receptor kinases: Crucial regulators of blood pressure. J Am Heart Assoc 5: e003519. doi: 10.1161/JAHA.116.003519
|
| [8] |
Bar-Shavit R, Maoz M, Kancharla A, et al. (2016) G protein-coupled receptors in cancer. Int J Mol Sci 17: 1320. doi: 10.3390/ijms17081320
|
| [9] |
Flower DR (1999) Modelling G-protein-coupled receptors for drug design. Biochim Biophys Acta 1422: 207–234. doi: 10.1016/S0304-4157(99)00006-4
|
| [10] |
Katritch V, Cherezov V, Stevens RC (2012) Diversity and modularity of G protein-coupled receptor structures. Trends Pharmacol Sci 33: 17–27. doi: 10.1016/j.tips.2011.09.003
|
| [11] | Gether U, Ballesteros JA, Seifert R, et al. (1997) Structural instability of a constitutively active G protein-coupled receptor. Agonist-independent activation due to conformational flexibility. J Biol Chem 272: 2587–2590. |
| [12] |
Seifert R, Wenzel-Seifert K (2002) Constitutive activity of G-protein-coupled receptors: cause of disease and common property of wild-type receptors. Naunyn Schmiedebergs Arch Pharmacol 366: 381–416. doi: 10.1007/s00210-002-0588-0
|
| [13] |
Nelson G, Hoon MA, Chandrashekar J, et al. (2001) Mammalian sweet taste receptors. Cell 106: 381–390. doi: 10.1016/S0092-8674(01)00451-2
|
| [14] |
Nelson G, Chandrashekar J, Hoon MA, et al. (2002) An amino-acid taste receptor. Nature 416: 199–202. doi: 10.1038/nature726
|
| [15] | Milligan G (2004) G protein-coupled receptor dimerization: Function and ligand pharmacology. Mol Pharmacol 66. |
| [16] |
Syrovatkina V, Alegre KO, Dey R, et al. (2016) Regulation, signaling, and physiological functions of G-proteins. J Mol Biol 428: 3850–3868. doi: 10.1016/j.jmb.2016.08.002
|
| [17] | Wess J (1997) G-protein-coupled receptors: molecular mechanisms involved in receptor activation and selectivity of G-protein recognition. FASEB J 11: 346–354. |
| [18] |
Rasumussen SG, DeVree BT, Zou Y, et al. (2011) Crystal structure of the beta2 adrenergic receptor-Gs protein complex. Nature 477: 549–555. doi: 10.1038/nature10361
|
| [19] |
Gilman AG (1987) G proteins: transducers of receptor-generated signals. Annu Rev Biochem 56: 615–649. doi: 10.1146/annurev.bi.56.070187.003151
|
| [20] | Nie J, Lewis DL (2001) Structural domains of the CB1 cannabinoid receptor that contribute to constitutive activity and G-protein sequestration. J Neurosci 21: 8758–8764. |
| [21] |
Kobilka BK, Deupi X (2007) Conformational complexity of G-protein-coupled receptors. Trends Pharmacol Sci 28: 397–406. doi: 10.1016/j.tips.2007.06.003
|
| [22] |
Ross EM, Wilkie TM (2000) GTPase-activating proteins for heterotrimeric G proteins: regulators of G protein signaling (RGS) and RGS-like proteins. Annu Rev Biochem 69: 795–827. doi: 10.1146/annurev.biochem.69.1.795
|
| [23] |
Claing A, Laporte SA, Caron MG, et al. (2002) Endocytosis of G protein-coupled receptors: roles of G protein-coupled receptor kinases and beta-arrestin proteins. Prog Neurobiol 66: 61–79. doi: 10.1016/S0301-0082(01)00023-5
|
| [24] |
Okada T, Le TI, Fox BA, et al. (2000) X-Ray diffraction analysis of three-dimensional crystals of bovine rhodopsin obtained from mixed micelles. J Struct Biol 130: 73–80. doi: 10.1006/jsbi.1999.4209
|
| [25] |
Palczewski K, Kumasaka T, Hori T, et al. (2000) Crystal structure of rhodopsin: A G protein-coupled receptor. Science 289: 739–745. doi: 10.1126/science.289.5480.739
|
| [26] |
Cherezov V, Rosenbaum DM, Hanson MA, et al. (2007) High resolution crystal structure of an engineered human beta2-adrenergic G protein-couple receptor. Science 318: 1258–1265. doi: 10.1126/science.1150577
|
| [27] |
Rasmussen SG, Choi HJ, Rosenbaum DM, et al. (2007) Crystal structure of the human beta2 adrenergic G-protein-coupled receptor. Nature 450: 383–387. doi: 10.1038/nature06325
|
| [28] | Stroud RM (2011) New tools in membrane protein determination. F1000 Biol Rep 3: 8. |
| [29] |
Ujwal R, Bowie JU (2011) Crystallizing membrane proteins using lipidic bicelles. Methods 55: 337–341. doi: 10.1016/j.ymeth.2011.09.020
|
| [30] |
Denisov IG, Sligar SG (2016) Nanodiscs for structural and functional studies of membrane proteins. Nat Struct Mol Biol 23: 481–486. doi: 10.1038/nsmb.3195
|
| [31] |
Xiang J, Chun E, Liu C, et al. (2016) Successful strategies to determine high-resolution structures of GPCRs. Trends Pharmacol Sci 37: 1055–1069. doi: 10.1016/j.tips.2016.09.009
|
| [32] |
Rawlings AE (2016) Membrane proteins: always an insoluble problem? Biochem Soc Trans 44: 790–795. doi: 10.1042/BST20160025
|
| [33] |
Neubig RR, Siderovski DP (2002) Regulators of G-protein signalling as new central nervous system drug targets. Nat Rev Drug Discov 1: 187–197. doi: 10.1038/nrd747
|
| [34] |
Bayburt TH, Grinkova YV, Sligar SG (2002) Self-assembly of discoidal phospholipid bilayer nanoparticles with membrane scaffold proteins. Nano Lett 2: 853–856. doi: 10.1021/nl025623k
|
| [35] |
Delmar JA, Bolla JR, Su CC, et al. (2015) Crystallization of membrane proteins by vapor diffusion. Methods Enzymol 557: 363–392. doi: 10.1016/bs.mie.2014.12.018
|
| [36] | Jaakola VP, Griffith MT, Hanson MA, et al. (2008) The 2.6 angstrom crystal structure of a human A2A adenosine receptor bound to an antagonist. Science 322: 1211–1217. |
| [37] |
Dore AS, Okrasa K, Patel JC, et al. (2014) Structure of class C GPCR metabotropic glutamate receptor 5 transmembrane domain. Nature 511: 557–562. doi: 10.1038/nature13396
|
| [38] |
Thompson AA, Liu W, Chun E, et al. (2012) Structure of the nociceptin/orphanin FQ receptor in complex with a peptide mimetic. Nature 485: 395–399. doi: 10.1038/nature11085
|
| [39] |
Wang C, Wu H, Katritch V, et al. (2013) Structure of the human smoothened receptor bound to an antitumour agent. Nature 497: 338–343. doi: 10.1038/nature12167
|
| [40] |
Yin J, Babaoglu K, Brautigam CA, et al. (2016) Structure and ligand-binding mechanism of the human OX1 and OX2 orexin receptors. Nat Struct Mol Biol 23: 293–299. doi: 10.1038/nsmb.3183
|
| [41] | Zhang K, Zhang J, Gao Z, et al. (2014) Structure of the human P2Y12 receptor in complex with an antithrombotic drug. Nature 509. |
| [42] | Isberg V, Mordalski S, Munk C, et al. (2016) GPCRdb: an information system for G protein-coupled receptors. Nucleic Acids Res. |
| [43] |
Berman HM, Westbrook J, Feng Z, et al. (2000) The protein data bank. Nucl Acids Res 28: 235–242. doi: 10.1093/nar/28.1.235
|
| [44] |
Teller DC, Okada T, Behnke CA, et al. (2001) Advances in determination of a high-resolution three-dimensional structure of rhodopsin, a model of G-protein-coupled receptors (GPCRs). Biochemistry 40: 7761–7772. doi: 10.1021/bi0155091
|
| [45] |
Yeagle PL, Choi G, Albert AD (2001) Studies on the structure of the G-protein-coupled receptor rhodopsin including the putative G-protein binding site in unactivated and activated forms. Biochemistry 40: 11932–11937. doi: 10.1021/bi015543f
|
| [46] |
Okada T, Fujiyoshi Y, Silow M, et al. (2002) Functional role of internal water molecules in rhodopsin revealed by X-ray crystallography. Proc Natl Acad Sci USA 99: 5982–5987. doi: 10.1073/pnas.082666399
|
| [47] |
Choi G, Landin J, Galan JF, et al. (2002) Structural studies of metarhodopsin II, the activated form of the G-protein coupled receptor, rhodopsin. Biochemistry 41: 7318–7324. doi: 10.1021/bi025507w
|
| [48] |
Li J, Edwards PC, Burghammer M, et al. (2004) Structure of bovine rhodopsin in a trigonal crystal form. J Mol Biol 343: 1409–1438. doi: 10.1016/j.jmb.2004.08.090
|
| [49] |
Okada T, Sugihara M, Bondar AN, et al. (2004) The retinal conformation and its environment in rhodopsin in light of a new 22 A crystal structure. J Mol Biol 342: 571–583. doi: 10.1016/j.jmb.2004.07.044
|
| [50] |
Nakamichi H, Okada T (2006) Crystallographic analysis of primary visual photochemistry. Angew Chem Int Ed Engl 45: 4270–4273. doi: 10.1002/anie.200600595
|
| [51] |
Nakamichi H, Okada T (2006) Local peptide movement in the photoreaction intermediate of rhodopsin. Proc Natl Acad Sci USA 103: 12729–12734. doi: 10.1073/pnas.0601765103
|
| [52] |
Salom D, Lodowski DT, Stenkamp RE, et al. (2006) Crystal structure of a photoactivated deprotonated intermediate of rhodopsin. Proc Natl Acad Sci USA 103: 16123–16128. doi: 10.1073/pnas.0608022103
|
| [53] |
Standfuss J, Xie G, Edwards PC, et al. (2007) Crystal structure of a thermally stable rhodopsin mutant. J Mol Biol 372: 1179–1188. doi: 10.1016/j.jmb.2007.03.007
|
| [54] |
Nakamichi H, Buss V, Okada T (2007) Photoisomerization mechanism of rhodopsin and 9-cis-rhodopsin revealed by x-ray crystallography. Biophys J 92: L106–L108. doi: 10.1529/biophysj.107.108225
|
| [55] | Stenkamp RE (2008) Alternative models for two crystal structures of bovine rhodopsin. Acta Crystallogr D D64: 902–904. |
| [56] |
Park JH, Scheerer P, Hofmann KP, et al. (2008) Crystal structure of the ligand-free G-protein-coupled receptor opsin. Nature 454: 183–187. doi: 10.1038/nature07063
|
| [57] |
Scheerer P, Park JH, Hildebrand PW, et al. (2008) Crystal structure of opsin in its G-protein-interacting conformation. Nature 455: 497–502. doi: 10.1038/nature07330
|
| [58] |
Standfuss J, Edwards PC, D'Antona A, et al. (2011) The structural basis of agonist-induced activation in constitutively active rhodopsin. Nature 471: 656–660. doi: 10.1038/nature09795
|
| [59] |
Makino CL, Riley CK, Looney J, et al. (2010) Binding of more than one retinoid to visual opsins. Biophys J 99: 2366–2373. doi: 10.1016/j.bpj.2010.08.003
|
| [60] |
Choe HW, Kim YJ, Park JH, et al. (2011) Crystal structure of metarhodopsin II. Nature 471: 651–655. doi: 10.1038/nature09789
|
| [61] |
Deupi X, Edwards P, Singhal A, et al. (2012) Stabilized G protein binding site in the structure of constitutively active metarhodopsin-II. Proc Natl Acad Sci USA 109: 119–124. doi: 10.1073/pnas.1114089108
|
| [62] |
Park JH, Morizumi T, Li Y, et al. (2013) Opsin, a structural model for olfactory receptors? Angew Chem Int Ed Engl 52: 11021–11024. doi: 10.1002/anie.201302374
|
| [63] |
Singhal A, Ostermaier MK, Vishnivetskiy SA, et al. (2013) Insights into congenital stationary night blindness based on the structure of G90D rhodopsin. EMBO Rep 14: 520–526. doi: 10.1038/embor.2013.44
|
| [64] |
Szczepek M, Beyriere F, Hofmann KP, et al. (2014) Crystal structure of a common GPCR-binding interface for G protein and arrestin. Nat Commun 5: 4801. doi: 10.1038/ncomms5801
|
| [65] |
Blankenship E, Vahedi-Faridi A, Lodowski DT (2015) The high-resolution structure of activated opsin reveals a conserved solvent network in the transmembrane region essential for activation. Structure 23: 2358–2364. doi: 10.1016/j.str.2015.09.015
|
| [66] |
Kang Y, Zhou XE, Gao X, et al. (2015) Crystal structure of rhodopsin bound to arrestin by femtosecond X-ray laser. Nature 523: 561–567. doi: 10.1038/nature14656
|
| [67] |
Singhal A, Guo Y, Matkovic M, et al. (2016) Structural role of the T94I rhodopsin mutation in congenital stationary night blindness. EMBO Rep 17: 1431–1440. doi: 10.15252/embr.201642671
|
| [68] |
Gulati S, Jastrzebska B, Banerjee S, et al. (2017) Photocyclic behavior of rhodopsin induced by an atypical isomerization mechanism. Proc Natl Acad Sci USA 114: E2608–E2615. doi: 10.1073/pnas.1617446114
|
| [69] |
Warne T, Serrano-Vega MJ, Baker JG, et al. (2008) Structure of a beta1-adrenergic G-protein-coupled receptor. Nature 454: 486–491. doi: 10.1038/nature07101
|
| [70] |
Warne T, Moukhametzianov R, Baker JG, et al. (2011) The structural basis for agonist and partial agonist action on a beta(1)-adrenergic receptor. Nature 469: 241–244. doi: 10.1038/nature09746
|
| [71] |
Moukhametzianov R, Warne T, Edwards PC, et al. (2011) Two distinct conformations of helix 6 observed in antagonist-bound structures of a beta-1 adrenergic receptor. Proc Natl Acad Sci USA 108: 8228–8232. doi: 10.1073/pnas.1100185108
|
| [72] |
Christopher JA, Brown J, Dore AS, et al. (2013) Biophysical fragment screening of the beta1-adrenergic receptor: identification of high affinity arylpiperazine leads using structure-based drug design. J Med Chem 56: 3446–3455. doi: 10.1021/jm400140q
|
| [73] |
Warne T, Edwards PC, Leslie AG, et al. (2012) Crystal structures of a stabilized beta1-adrenoceptor bound to the biased agonists bucindolol and carvedilol. Structure 20: 841–849. doi: 10.1016/j.str.2012.03.014
|
| [74] | Miller-Gallacher JL, Nehme R, Warne T, et al. (2014) The 2.1 A resolution structure of cyanopindolol-bound beta1-adrenoceptor identifies an intramembrane Na+ ion that stabilises the ligand-free receptor. PLoS One 9: e92727. |
| [75] |
Huang J, Chen S, Zhang JJ, et al. (2013) Crystal structure of oligomeric beta1-adrenergic G protein-coupled receptors in ligand-free basal state. Nat Struct Mol Biol 20: 419–425. doi: 10.1038/nsmb.2504
|
| [76] |
Sato T, Baker J, Warne T, et al. (2015) Pharmacological analysis and structure determination of 7-Methylcyanopindolol-bound beta1-adrenergic receptor. Mol Pharmacol 88: 1024–1034. doi: 10.1124/mol.115.101030
|
| [77] |
Leslie AG, Warne T, Tate CG (2015) Ligand occupancy in crystal structure of beta1-adrenergic G protein-coupled receptor. Nat Struct Mol Biol 22: 941–942. doi: 10.1038/nsmb.3130
|
| [78] | Hanson MA, Cherezov V, Griffith MT, et al. (2008) A specific cholesterol binding site is established by the 2.8 A structure of the human beta2-adrenergic receptor. Structure 6: 897–905. |
| [79] |
Bokoch MP, Zou Y, Rasmussen SG, et al. (2010) Ligand-specific regulation of the extracellular surface of a G-protein-coupled receptor. Nature 463: 108–112. doi: 10.1038/nature08650
|
| [80] |
Wacker D, Fenalti G, Brown MA, et al. (2010) Conserved binding mode of human beta2 adrenergic receptor inverse agonists and antagonist revealed by X-ray crystallography. J Am Chem Soc 132: 11443–11445. doi: 10.1021/ja105108q
|
| [81] |
Rasmussen SG, Choi HJ, Fung JJ, et al. (2011) Structure of a nanobody-stabilized active state of the beta(2) adrenoceptor. Nature 469: 175–180. doi: 10.1038/nature09648
|
| [82] |
Rosenbaum DM, Zhang C, Lyons JA, et al. (2011) Structure and function of an irreversible agonist-beta(2) adrenoceptor complex. Nature 469: 236–240. doi: 10.1038/nature09665
|
| [83] | Zou Y, Weis WI, Kobilka BK (2012) N-terminal T4 lysozyme fusion facilitates crystallization of a G protein coupled receptor. PLos One 7. |
| [84] |
Ring AM, Manglik A, Kruse AC, et al. (2013) Adrenaline-activated structure of beta2-adrenoreceptor stabilized by an engineered nanobody. Nature 502: 575–579. doi: 10.1038/nature12572
|
| [85] |
Weichert D, Kruse AC, Manglik A, et al. (2014) Covalent agonists for studying G protein-coupled receptor activation. Proc Natl Acad Sci USA 111: 10744–10748. doi: 10.1073/pnas.1410415111
|
| [86] |
Huang CY, Olieric V, Ma P, et al. (2016) In meso in situ serial X-ray crystallography of soluble and membrane proteins at cryogenic temperatures. Acta Crystallogr D 72: 93–112. doi: 10.1107/S2059798315021683
|
| [87] |
Staus DP, Strachan RT, Manglik A, et al. (2016) Allosteric nanobodies reveal the dynamic range and diverse mechanisms of G-protein-coupled receptor activation. Nature 535: 448–452. doi: 10.1038/nature18636
|
| [88] |
Shimamura T, Shiroishi M, Weyand S, et al. (2011) Structure of the human histamine H1 receptor complex with doxepin. Nature 475: 65–70. doi: 10.1038/nature10236
|
| [89] |
Wang C, Jiang Y, Ma J, et al. (2013) Structural basis for molecular recognition at serotonin receptors. Science 340: 610–614. doi: 10.1126/science.1232807
|
| [90] |
Wacker D, Wang C, Katritch V, et al. (2013) Structural features for functional selectivity at serotonin receptors. Science 340: 615–619. doi: 10.1126/science.1232808
|
| [91] |
Liu W, Wacker D, Gati C, et al. (2013) Serial femtosecond crystallography of G protein-coupled receptors. Science 342: 1521–1524. doi: 10.1126/science.1244142
|
| [92] |
Wacker D, Wang S, McCorvy JD, et al. (2017) Crystal structure of an LSD-bound human serotonin receptor. Cell 168: 377–389. doi: 10.1016/j.cell.2016.12.033
|
| [93] |
Thal DM, Sun B, Feng D, et al. (2016) Crystal structures of the M1 and M4 muscarinic acetylcholine receptors. Nature 531: 335–340. doi: 10.1038/nature17188
|
| [94] |
Haga K, Kruse AC, Asada H, et al. (2012) Structure of the human M2 muscarinic acetylcholine receptor bound to an antagonist. Nature 482: 547–551. doi: 10.1038/nature10753
|
| [95] |
Kruse AC, Ring AM, Manglik A, et al. (2013) Activation and allosteric modulation of a muscarinic acetylcholine receptor. Nature 504: 101–106. doi: 10.1038/nature12735
|
| [96] |
Kruse AC, Hu J, Pan AC, et al. (2012) Structure and dynamics of the M3 muscarinic acetylcholine receptor. Nature 482: 552–556. doi: 10.1038/nature10867
|
| [97] |
Thorsen TS, Matt R, Weis WI, et al. (2014) Modified T4 lysozyme fusion proteins facilitate G protein-coupled receptor crystallogenesis. Structure 22: 1657–1664. doi: 10.1016/j.str.2014.08.022
|
| [98] |
Glukhova A, Thal DM, Nguyen AT, et al. (2017) Structure of the adenosine A1 receptor reveals the basis for subtype selectivity. Cell 168: 867–877. doi: 10.1016/j.cell.2017.01.042
|
| [99] |
Dore AS, Robertson N, Errey JC, et al. (2011) Structure of the adenosine A(2A) receptor in complex with ZM241385 and the xanthines XAC and caffeine. Structure 19: 1283–1293. doi: 10.1016/j.str.2011.06.014
|
| [100] |
Xu F, Wu H, Katritch V, et al. (2011) Structure of an agonist-bound human A2A adenosine receptor. Science 332: 322–327. doi: 10.1126/science.1202793
|
| [101] |
Lebon G, Warne T, Edwards PC, et al. (2011) Agonist-bound adenosine A2A receptor structures reveal common features of GPCR activation. Nature 474: 521–525. doi: 10.1038/nature10136
|
| [102] | Hino T, Arakawa T, Iwanari H, et al. (2012) G-protein-coupled receptor inactivation by an allosteric inverse-agonist antibody. Nature 482: 237–240. |
| [103] |
Congreve M, Andrews SP, Dore AS, et al. (2012) Discovery of 1,2,4-triazine derivatives as adenosine A(2A) antagonists using structure based drug design. J Med Chem 55: 1898–1903. doi: 10.1021/jm201376w
|
| [104] |
Liu W, Chun E, Thompson AA, et al. (2012) Structural basis for allosteric regulation of GPCRs by sodium ions. Science 337: 232–236. doi: 10.1126/science.1219218
|
| [105] |
Lebon G, Edwards PC, Leslie AG, et al. (2015) Molecular determinants of CGS21680 binding to the human adenosine A2A receptor. Mol Pharmacol 87: 907–915. doi: 10.1124/mol.114.097360
|
| [106] |
Segala E, Guo D, Cheng RK, et al. (2016) Controlling the dissociation of ligands from the adenosine A2A receptor through modulation of salt bridge strength. J Med Chem 59: 6470–6479. doi: 10.1021/acs.jmedchem.6b00653
|
| [107] |
Batyuk A, Galli L, Ishchenko A, et al. (2016) Native phasing of x-ray free-electron laser data for a G protein-coupled receptor. Sci Adv 2: e1600292. doi: 10.1126/sciadv.1600292
|
| [108] |
Carpenter B, Nehme R, Warne T, et al. (2016) Structure of the adenosine A(2A) receptor bound to an engineered G protein. Nature 536: 104–107. doi: 10.1038/nature18966
|
| [109] |
Sun B, Bachhawat P, Chu ML, et al. (2017) Crystal structure of the adenosine A2A receptor bound to an antagonist reveals a potential allosteric pocket. Proc Natl Acad Sci USA 114: 2066–2071. doi: 10.1073/pnas.1621423114
|
| [110] |
Hanson MA, Roth CB, Jo E, et al. (2012) Crystal structure of a lipid G protein-coupled receptor. Science 335: 851–855. doi: 10.1126/science.1215904
|
| [111] |
Chrencik JE, Roth CB, Terakado M, et al. (2015) Crystal structure of antagonist bound human lysophosphatidic acid receptor 1. Cell 161: 1633–1643. doi: 10.1016/j.cell.2015.06.002
|
| [112] |
Hua T, Vemuri K, Pu M, et al. (2016) Crystal structure of the human cannabinoid receptor CB1. Cell 167: 750–762. doi: 10.1016/j.cell.2016.10.004
|
| [113] | Shao Z, Yin J, Chapman K, et al. (2016) High-resolution crystal structure of the human CB1 cannabinoid receptor. Nature. |
| [114] |
White JF, Noinaj N, Shibata Y, et al. (2012) Structure of the agonist-bound neurotensin receptor. Nature 490: 508–513. doi: 10.1038/nature11558
|
| [115] |
Egloff P, Hillenbrand M, Klenk C, et al. (2014) Structure of signaling-competent neurotensin receptor 1 obtained by directed evolution in Escherichia coli. Proc Natl Acad Sci USA 111: E655–E662. doi: 10.1073/pnas.1317903111
|
| [116] |
Krumm BE, White JF, Shah P, et al. (2015) Structural prerequisites for G-protein activation by the neurotensin receptor. Nat Commun 6: 7895. doi: 10.1038/ncomms8895
|
| [117] |
Krumm BE, Lee S, Bhattacharya S, et al. (2016) Structure and dynamics of a constitutively active neurotensin receptor. Sci Rep 6: 38564. doi: 10.1038/srep38564
|
| [118] | Yin J, Mobarec JC, Kolb P, et al. (2015) Crystal structure of the human OX2 orexin receptor bound to the insomnia drug suvorexant. Nature 519: 247–250. |
| [119] |
Gayen A, Goswami SK, Mukhopadhyay C (2011) NMR evidence of GM1-induced conformational change of Substance P using isotropic bicelles. Biochim Biophys Acta 1808: 127–139. doi: 10.1016/j.bbamem.2010.09.023
|
| [120] |
Shihoya W, Nishizawa T, Okuta A, et al. (2016) Activation mechanism of endothelin ETB receptor by endothelin-1. Nature 537: 363–368. doi: 10.1038/nature19319
|
| [121] | Park SH, Das BB, Casagrande F, et al. (2012) Structure of the chemokine receptor CXCR1 in phospholipid bilayers. Nature 491: 779–783. |
| [122] |
Zheng Y, Qin L, Zacarias NV, et al. (2016) Structure of CC chemokine receptor 2 with orthosteric and allosteric antagonists. Nature 540: 458–461. doi: 10.1038/nature20605
|
| [123] |
Wu B, Chien EY, Mol CD, et al. (2010) Structures of the CXCR4 chemokine GPCR with small-molecule and cyclic peptide antagonists. Science 330: 1066–1071. doi: 10.1126/science.1194396
|
| [124] | Qin L, Kufareva I, Holden LG, et al. (2015) Structural biology. Crystal structure of the chemokine receptor CXCR4 in complex with a viral chemokine. Science 347: 1117–1122. |
| [125] |
Tan Q, Zhu Y, Li J, et al. (2013) Structure of the CCR5 chemokine receptor-HIV entry inhibitor maraviroc complex. Science 341: 1387–1390. doi: 10.1126/science.1241475
|
| [126] |
Oswald C, Rappas M, Kean J, et al. (2016) Intracellular allosteric antagonism of the CCR9 receptor. Nature 540: 462–465. doi: 10.1038/nature20606
|
| [127] |
Miller RL, Thompson AA, Trapella C, et al. (2015) The importance of ligand-receptor conformational pairs in stabilization: Spotlight on the N/OFQ G protein-coupled receptor. Structure 23: 2291–2299. doi: 10.1016/j.str.2015.07.024
|
| [128] |
Wu H, Wacker D, Mileni M, et al. (2012) Structure of the human kappa-opioid receptor in complex with JDTic. Nature 485: 327–332. doi: 10.1038/nature10939
|
| [129] |
Manglik A, Kruse AC, Kobilka TS, et al. (2012) Crystal structure of the mu-opioid receptor bound to a morphinan antagonist. Nature 485: 321–326. doi: 10.1038/nature10954
|
| [130] |
Huang W, Manglik A, Venkatakrishnan AJ, et al. (2015) Structural insights into micro-opioid receptor activation. Nature 524: 315–321. doi: 10.1038/nature14886
|
| [131] |
Granier S, Manglik A, Kruse AC, et al. (2012) Structure of the delta-opioid receptor bound to naltrindole. Nature 485: 400–404. doi: 10.1038/nature11111
|
| [132] |
Fenalti G, Giguere PM, Katritch V, et al. (2014) Molecular control of delta-opioid receptor signaling. Nature 506: 191–196. doi: 10.1038/nature12944
|
| [133] |
Fenalti G, Zatsepin NA, Betti C, et al. (2015) Structural basis for bifunctional peptide recognition at human delta-opioid receptor. Nat Struct Mol Biol 22: 265–268. doi: 10.1038/nsmb.2965
|
| [134] |
Zhang H, Unal H, Gati C, et al. (2015) Structure of the Angiotensin receptor revealed by serial femtosecond crystallography. Cell 161: 833–844. doi: 10.1016/j.cell.2015.04.011
|
| [135] |
Zhang H, Unal H, Desnoyer R, et al. (2015) Structural basis for ligand recognition and functional selectivity at angiotensin receptor. J Biol Chem 290: 29127–29139. doi: 10.1074/jbc.M115.689000
|
| [136] | Zhang H, Han GW, Batyuk A, et al. (2017) Structural basis for selectivity and diversity in angiotensin II receptors. Nature. |
| [137] | Burg JS, Ingram JR, Venkatakrishnan AJ, et al. (2015) Structural biology. Structural basis for chemokine recognition and activation of a viral G protein-coupled receptor. Science 347: 1113–1117. |
| [138] |
Zhang D, Gao ZG, Zhang K, et al. (2015) Two disparate ligand-binding sites in the human P2Y1 receptor. Nature 520: 317–321. doi: 10.1038/nature14287
|
| [139] |
Zhang J, Zhang K, Gao ZG, et al. (2014) Agonist-bound structure of the human P2Y12 receptor. Nature 509: 119–122. doi: 10.1038/nature13288
|
| [140] |
Zhang C, Srinivasan Y, Arlow DH, et al. (2012) High-resolution crystal structure of human protease-activated receptor 1. Nature 492: 387–392. doi: 10.1038/nature11701
|
| [141] |
Srivastava A, Yano J, Hirozane Y, et al. (2014) High-resolution structure of the human GPR40 receptor bound to allosteric agonist TAK-875. Nature 513: 124–127. doi: 10.1038/nature13494
|
| [142] |
Siu FY, He M, de Graaf C, et al. (2013) Structure of the human glucagon class B G-protein-coupled receptor. Nature 499: 444–449. doi: 10.1038/nature12393
|
| [143] |
Jazayeri A, Dore AS, Lamb D, et al. (2016) Extra-helical binding site of a glucagon receptor antagonist. Nature 533: 274–277. doi: 10.1038/nature17414
|
| [144] |
Hollenstein K, Kean J, Bortolato A, et al. (2013) Structure of class B GPCR corticotropin-releasing factor receptor 1. Nature 499: 438–443. doi: 10.1038/nature12357
|
| [145] | Dore AS, Bortolato A, Hollenstein K, et al. (2017) Decoding corticotropin-releasing factor receptor type 1 crystal structures. Curr Mol Pharmacol: Epub ahead of print, DOI: 10.2174/1874467210666170110114727. |
| [146] |
Wu H, Wang C, Gregory KJ, et al. (2014) Structure of a class C GPCR metabotropic glutamate receptor 1 bound to an allosteric modulator. Science 344: 58–64. doi: 10.1126/science.1249489
|
| [147] |
Christopher JA, Aves SJ, Bennett KA, et al. (2015) Fragment and structure-based drug discovery for a class C GPCR: Discovery of the mGlu5 negative allosteric modulator HTL14242 (3-Chloro-5-[6-(5-fluoropyridin-2-yl)pyrimidin-4-yl]benzonitrile). J Med Chem 58: 6653–6664. doi: 10.1021/acs.jmedchem.5b00892
|
| [148] | Wang C, Wu H, Evron T, et al. (2014) Structural basis for Smoothened receptor modulation and chemoresistance to anticancer drugs. Nat Commun 5: 4355. |
| [149] |
Byrne EF, Sircar R, Miller PS, et al. (2016) Structural basis of Smoothened regulation by its extracellular domains. Nature 535: 517–522. doi: 10.1038/nature18934
|
| [150] |
Lundstrom K (2006) Latest development in drug discovery on G protein-coupled receptors. Curr Protein Pept Sci 7: 465–470. doi: 10.2174/138920306778559403
|
| [151] |
Shonberg J, Kling RC, Gmeiner P, et al. (2015) GPCR crystal structures: Medicinal chemistry in the pocket. Bioorg Med Chem 23: 3880–3906. doi: 10.1016/j.bmc.2014.12.034
|
| [152] |
Attwood TK, Findlay JB (1993) Design of a discriminating fingerprint for G-protein-coupled receptors. Protein Eng 6: 167–176. doi: 10.1093/protein/6.2.167
|
| [153] |
Chee MS, Satchwell SC, Preddie E, et al. (1990) Human cytomegalovirus encodes three G protein-coupled receptor homologues. Nature 344: 774–777. doi: 10.1038/344774a0
|
| [154] |
Attwood TK, Findlay JB (1994) Fingerprinting G-protein-coupled receptors. Protein Eng 7: 195–203. doi: 10.1093/protein/7.2.195
|
| [155] | Strader CD, Sigal IS, Dixon RA (1989) Structural basis of beta-adrenergic receptor function. FASEB J 3: 1825–1832. |
| [156] | Liapakis G, Ballesteros JA, Papachristou S, et al. (2000) The forgotten serine. A critical role for Ser-2035.42 in ligand binding to and activation of the beta 2-adrenergic receptor. J Biol Chem 275: 37779–37788. |
| [157] | Swaminath G, Xiang Y, Lee TW, et al. (2004) Sequential binding of agonists to the beta2 adrenoceptor. Kinetic evidence for intermediate conformational states. J Biol Chem 279: 686–691. |
| [158] |
Baldwin JM (1994) Structure and function of receptors coupled to G proteins. Curr Opin Cell Biol 6: 180–190. doi: 10.1016/0955-0674(94)90134-1
|
| [159] |
Tyndall JD, Sandilya R (2005) GPCR agonists and antagonists in the clinic. Med Chem 1: 405–421. doi: 10.2174/1573406054368675
|
| [160] | Jacoby E, Bouhelal R, Gerspacher M, et al. (2006) The 7 TM G-protein-coupled receptor target family. Chem Med Chem 1: 761–782. |
| [161] | Spiss CK, Maze M (1985) Adrenoreceptors. Anaesthesist 34: 1–10. |
| [162] |
Civelli O, Reinscheid RK, Zhang Y, et al. (2013) G protein-coupled receptor deorphanizations. Annu Rev Pharmacol Toxicol 53: 127–146. doi: 10.1146/annurev-pharmtox-010611-134548
|
| [163] |
Barst RJ, Langleben D, Frost A, et al. (2004) Sitaxsentan therapy for pulmonary arterial hypertension. Am J Respir Crit Care Med 169: 441–447. doi: 10.1164/rccm.200307-957OC
|
| [164] |
Kotake T, Usami M, Akaza H, et al. (1999) Goserelin acetate with or without antiandrogen or estrogen in the treatment of patients with advanced prostate cancer: a multicenter, randomized, controlled trial in Japan. Zoladex Study Group. Jpn J Clin Oncol 29: 562–570. doi: 10.1093/jjco/29.11.562
|
| [165] |
Onuffer JJ, Horuk R (2002) Chemokines, chemokine receptors and small-molecule antagonists: recent developments. Trends Pharmacol Sci 23: 459–467. doi: 10.1016/S0165-6147(02)02064-3
|
| [166] |
Fatkenheuer G, Pozniak AL, Johnson MA, et al. (2005) Efficacy of short-term monotherapy with maraviroc, a new CCR5 antagonist, in patients infected with HIV-1. Nat Med 11: 1170–1172. doi: 10.1038/nm1319
|
| [167] |
Lu M, Wu B (2016) Structural studies of G protein-coupled receptors. IUBMB Life 68: 894–903. doi: 10.1002/iub.1578
|
| [168] |
Strotmann R, Schrock K, Boselt I, et al. (2011) Evolution of GPCR: change and continuity. Mol Cell Endocrinol 331: 170–178. doi: 10.1016/j.mce.2010.07.012
|
| [169] |
Ballesteros JA, Weinstein H (1995) Integrated methods for the construction of three-dimensional models and computational probing of structure-function relations in G protein-coupled receptors. Method Neurosci 25: 366–428. doi: 10.1016/S1043-9471(05)80049-7
|
| [170] |
Zhang D, Weinstein H (1994) Polarity conserved positions in transmembrane domains of G-protein coupled receptors and bacteriorhodopsin. FEBS Lett 337: 207–212. doi: 10.1016/0014-5793(94)80274-2
|
| [171] |
Katritch V, Fenalti G, Abola EE, et al. (2014) Allosteric sodium in class A GPCR signaling. Trends Biochem Sci 39: 233–244. doi: 10.1016/j.tibs.2014.03.002
|
| [172] |
Bjarnadottir TK, Geirardsdottir K, Ingemansson M, et al. (2007) Identification of novel splice variants of Adhesion G protein-coupled receptors. Gene 387: 38–48. doi: 10.1016/j.gene.2006.07.039
|
| [173] |
Lin HH, Chang GW, Davies JQ, et al. (2004) Autocatalytic cleavage of the EMR2 receptor occurs at a conserved G protein-coupled receptor proteolytic site motif. J Biol Chem 279: 31823–31832. doi: 10.1074/jbc.M402974200
|
| [174] |
Isberg V, Vroling B, Van DKR, et al. (2014) GPCRDB: an information system for G protein-coupled receptors. Nucleic Acids Res 42: D422–D425. doi: 10.1093/nar/gkt1255
|
| [175] |
Gasparini F, Kuhn R, Pin JP (2002) Allosteric modulators of group I metabotropic glutamate receptors: novel subtype-selective ligands and therapeutic perspectives. Curr Opin Pharmacol 2: 43–49. doi: 10.1016/S1471-4892(01)00119-9
|
| [176] |
Malherbe P, Kratochwil N, Zenner MT, et al. (2003) Mutational analysis and molecular modeling of the binding pocket of the metabotropic glutamate 5 receptor negative modulator 2-methyl-6-(phenylethynyl)-pyridine. Mol Pharmacol 64: 823–832. doi: 10.1124/mol.64.4.823
|
| [177] | Litschig S, Gasparini F, Rueegg D, et al. (1999) CPCCOEt, a noncompetitive metabotropic glutamate receptor 1 antagonist, inhibits receptor signaling without affecting glutamate binding. Mol Pharmacol 55: 453–461. |
| [178] |
Silve C, Petrel C, Leroy C, et al. (2005) Delineating a Ca2+ binding pocket within the venus flytrap module of the human calcium-sensing receptor. J Biol Chem 280: 37917–37923. doi: 10.1074/jbc.M506263200
|
| [179] |
Hermans E, Challiss RA (2001) Structural, signalling and regulatory properties of the group I metabotropic glutamate receptors: prototypic family C G-protein-coupled receptors. Biochem J 359: 465–484. doi: 10.1042/bj3590465
|
| [180] |
Nakanishi S (1992) Molecular diversity of glutamate receptors and implications for brain function. Science 258: 597–603. doi: 10.1126/science.1329206
|
| [181] |
Riedel G, Platt B, Micheau J (2003) Glutamate receptor function in learning and memory. Behav Brain Res 140: 1–47. doi: 10.1016/S0166-4328(02)00272-3
|
| [182] |
Kunishima N, Shimada Y, Tsuji Y, et al. (2000) Structural basis of glutamate recognition by a dimeric metabotropic glutamate receptor. Nature 407: 971–977. doi: 10.1038/35039564
|
| [183] |
Niswender CM, Conn PJ (2010) Metabotropic glutamate receptors: physiology, pharmacology, and disease. Annu Rev Pharmacol Toxicol 50: 295–322. doi: 10.1146/annurev.pharmtox.011008.145533
|
| [184] |
Bhanot P, Brink M, Samos CH, et al. (1996) A new member of the frizzled family from Drosophila functions as a Wingless receptor. Nature 382: 225–230. doi: 10.1038/382225a0
|
| [185] |
Murone M, Rosenthal A, de Sauvage FJ (1999) Sonic hedgehog signaling by the patched-smoothened receptor complex. Curr Biol 9: 76–84. doi: 10.1016/S0960-9822(99)80018-9
|
| [186] |
Chen CM, Strapps W, Tomlinson A, et al. (2004) Evidence that the cysteine-rich domain of Drosophila Frizzled family receptors is dispensable for transducing Wingless. Proc Natl Acad Sci USA 101: 15961–15966. doi: 10.1073/pnas.0407103101
|
| [187] |
Nakano Y, Nystedt S, Shivdasani AA, et al. (2004) Functional domains and sub-cellular distribution of the Hedgehog transducing protein Smoothened in Drosophila. Mech Dev 121: 507–518. doi: 10.1016/j.mod.2004.04.015
|
| [188] |
Isberg V, de Graaf C, Bortolato A, et al. (2015) Generic GPCR residue numbers-Aligning topology maps minding the gaps. Trends Pharmacol Sci 36: 22–31. doi: 10.1016/j.tips.2014.11.001
|
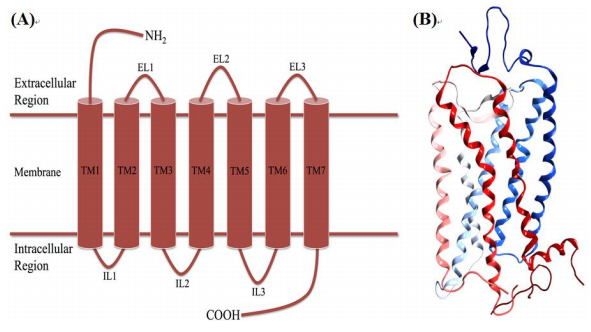
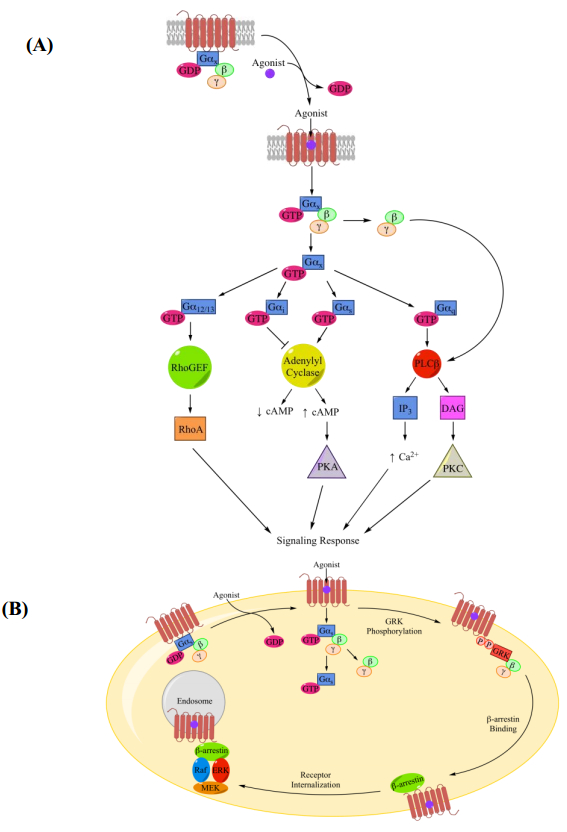
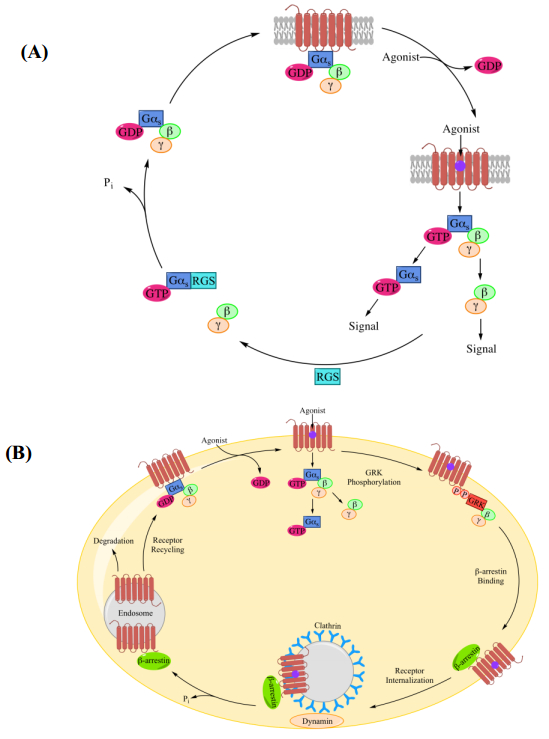
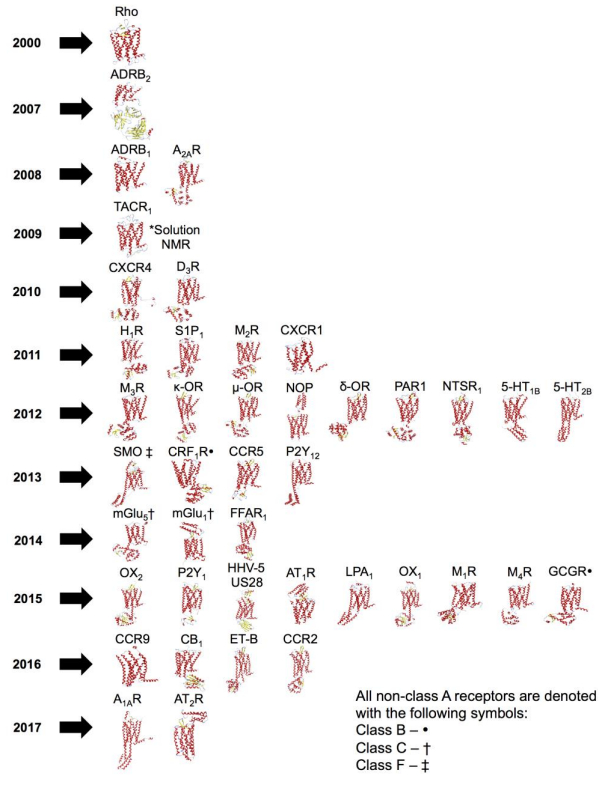
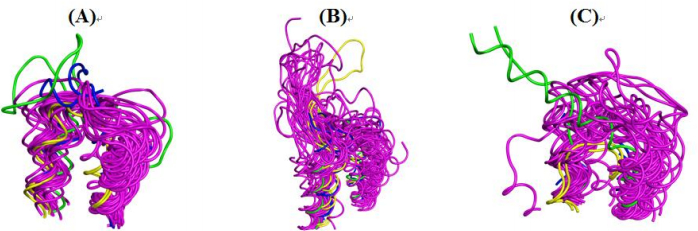
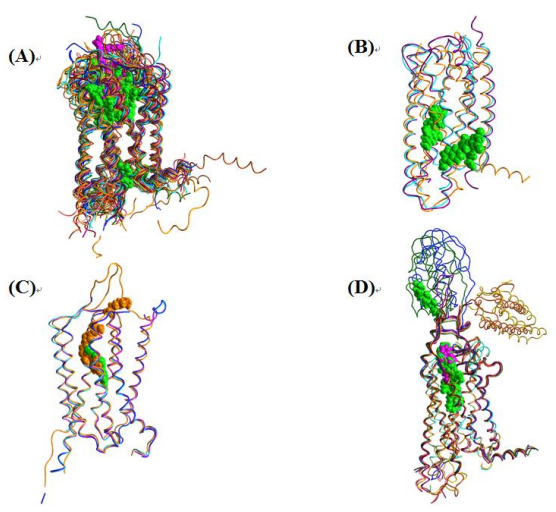
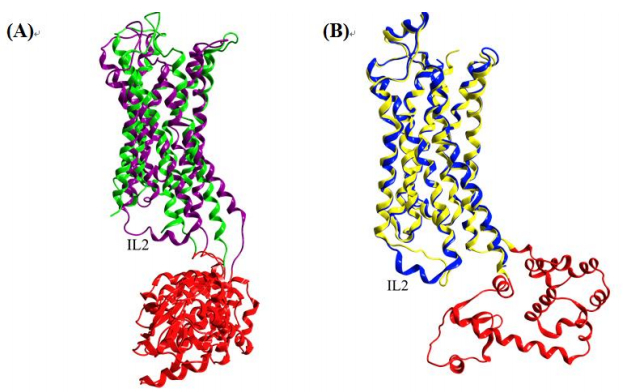
Figures(7) / Tables(2)

Samantha B. Gacasan, Daniel L. Baker, Abby L. Parrill. G protein-coupled receptors: the evolution of structural insight[J]. AIMS Biophysics, 2017, 4(3): 491-527. doi: 10.3934/biophy.2017.3.491







 DownLoad:
DownLoad: