Citation: Fabrizio Gentile, Alessia Arcaro, Stefania Pizzimenti, Martina Daga, Giovanni Paolo Cetrangolo, Chiara Dianzani, Alessio Lepore, Maria Graf, Paul R. J. Ames, Giuseppina Barrera. DNA damage by lipid peroxidation products: implications in cancer, inflammation and autoimmunity[J]. AIMS Genetics, 2017, 4(2): 103-137. doi: 10.3934/genet.2017.2.103

| [1] |
Csala M, Kardon T, Legeza B, et al. (2015) On the role of 4-hydroxynonenal in health and disease. Biochim Biophys Acta 1852: 826-838. doi: 10.1016/j.bbadis.2015.01.015

|
| [2] |
Forman HJ (2016) Redox signaling: An evolution from free radicals to aging. Free Radic Biol Med 97: 398-407. doi: 10.1016/j.freeradbiomed.2016.07.003
|
| [3] |
Hammer A, Ferro M, Tillian HM, et al. (1997) Effect of oxidative stress by iron on 4-hydroxynonenal formation and proliferative activity in hepatomas of different degrees of differentiation. Free Radic Biol Med 23: 26-33. doi: 10.1016/S0891-5849(96)00630-2
|
| [4] |
Canuto RA, Muzio G, Maggiora M, et al. (1993) Glutathione-S-transferase, alcohol dehydrogenase and aldehyde reductase activities during diethylnitrosamine-carcinogenesis in rat liver. Cancer Lett 68: 177-183. doi: 10.1016/0304-3835(93)90144-X
|
| [5] |
Barrera G, Pizzimenti S, Dianzani MU (2008) Lipid peroxidation: control of cell proliferation, cell differentiation and cell death. Mol Aspects Med 29: 1-8. doi: 10.1016/j.mam.2007.09.012
|
| [6] |
Voulgaridou GP, Anestopoulos I, Franco R, et al. (2011) DNA damage induced by endogenous aldehydes: Current state of knowledge. Mutat Res 711: 13-27. doi: 10.1016/j.mrfmmm.2011.03.006
|
| [7] |
Esterbauer H, Zollner H (1989) Methods for determination of aldehydic lipid peroxidation products. Free Radic Biol Med 7: 197-203. doi: 10.1016/0891-5849(89)90015-4
|
| [8] |
Frijhoff J, Winyard PG, Zarkovic N, et al. (2015) Clinical Relevance of Biomarkers of Oxidative Stress. Antioxid Redox Signal 23: 1144-1170. doi: 10.1089/ars.2015.6317
|
| [9] | Spiteller P, Kern W, Reiner J, et al. (2001) Aldehydic lipid peroxidation products derived from linoleic acid. Biochim Biophys Acta Mol Cell Biol L 1531: 188-208. |
| [10] |
Guéraud F, Atalay M, Bresgen N, et al. (2010) Chemistry and biochemistry of lipid peroxidation products. Free Radic Res 44: 1098-1124. doi: 10.3109/10715762.2010.498477
|
| [11] |
Esterbauer H, Schaur RJ, Zollner H (1991) Chemistry and biochemistry of 4-hydroxynonenal, malonaldehyde and related aldehydes. Free Radic Biol Med 11: 81-128 doi: 10.1016/0891-5849(91)90192-6
|
| [12] |
Kawai Y, Takeda S, Terao J (2007) Lipidomic analysis for lipid peroxidation-derived aldehydes using gas chromatography-mass spectrometry. Chem Res Toxicol 20: 99-107. doi: 10.1021/tx060199e
|
| [13] | Schauenstein E (1967) Autoxidation of polyunsaturated esters in water: chemical structure and biological activity of the products. J Lipid Res 8: 417-428. |
| [14] | Pryor WA, Porter NA (1990) Suggested mechanisms for the production of 4-hydroxy-2-nonenal from the autoxidation of polyunsaturated fatty acids. Free Radic Res Commun 8: 541-543. |
| [15] |
Benedetti A, Comporti M, Esterbauer H (1980) Identification of 4-hydroxynonenal as a cytotoxic product originating from the peroxidation of liver microsomal lipids. Biochim Biophys Acta 620: 281-296. doi: 10.1016/0005-2760(80)90209-X
|
| [16] |
Poli G, Dianzani MU, Cheeseman KH, et al. (1985) Separation and characterization of the aldehydic products of lipid peroxidation stimulated by carbon tetrachloride or ADP-iron in isolated rat hepatocytes and rat liver microsomal suspensions. Biochem J 227: 629-638. doi: 10.1042/bj2270629
|
| [17] |
Singh M, Kapoor A, Bhatnagar A (2015) Oxidative and reductive metabolism of lipid-peroxidation derived carbonyls. Chem Biol Interact 234: 261-273. doi: 10.1016/j.cbi.2014.12.028
|
| [18] | Barrera G, Pizzimenti S, Ciamporcero ES, et al. (2015) Role of 4-hydroxynonenal-protein adducts in human disease. Antiox Redox Signal 22: 18. |
| [19] |
Lee SH, Blair IA (2000) Characterization of 4-oxo-2-nonenal as a novel product of lipid peroxidation. Chem Res Toxicol 13: 698-702. doi: 10.1021/tx000101a
|
| [20] |
Doorn JA, Petersen DR (2002) Covalent modification of amino acid nucleophiles by the lipid peroxidation products 4-hydroxy-2-nonenal and 4-oxo-2-nonenal. Chem Res Toxicol 15: 1445-1450. doi: 10.1021/tx025590o
|
| [21] |
Rindgen D, Lee SH, Nakajima M, et al. (2000) Formation of a substituted 1,N6-etheno-2'-deoxyadenosine adduct by lipid hydroperoxide-mediated genration of 4-oxo-2-nonenal. Chem Res Toxicol 13: 846-852. doi: 10.1021/tx0000771
|
| [22] |
Lee SH, Rindgen D, Bible RH, et al. (2000) Characterization of 2'-deoxyadenosine adducts derived from 4-oxo-2-nonenal, a novel product of lipid peroxidation. Chem Res Toxicol 13: 565-574. doi: 10.1021/tx000057z
|
| [23] |
Pollack M, Oe T, Lee SH, et al. (2003) Characterization of 2'-deoxycytidine adducts derived from 4-oxo-2-nonenal, a novel lipid peroxidation product. Chem Res Toxicol 16: 893-900. doi: 10.1021/tx030009p
|
| [24] | Maekawa M, Kawai K, Takahashi Y, et al. (2005) Identification of 4-oxo-2-hexenal and other direct mutagens formed in model lipid peroxidation reactions as dGuo adducts. Chem Res Toxicol 19: 130-138. |
| [25] |
Uchida K, Kanematsu M, Sakai K, et al. (1998) Protein-bound acrolein: potential markers for oxidative stress. Proc Natl Acad Sci USA 95: 4882-4887. doi: 10.1073/pnas.95.9.4882
|
| [26] | Anderson MM, Hazen SL, Hsu FF, et al. (1997) Human neutrophils employ the myeloperoxidase-hydrogen peroxide-chloride system to convert hydroxy-amino acids into glycolaldehyde, 2-hydroxypropanal, and acrolein. A mechanism for the generation of highly reactive alpha-hydroxy and alpha, beta-unsaturated aldehydes by phagocytes at sites of inflammation. J Clin Invest 99: 424-432. |
| [27] |
Marnett LJ, Hurd HK, Hollstein MC, et al. (1985) Naturally occurring carbonyl compounds are mutagens in Salmonella tester strain TA104. Mutat Res 148: 25-34. doi: 10.1016/0027-5107(85)90204-0
|
| [28] |
Niedernhofer LJ, Daniels JS, Rouzer CA, et al. (2003) Malondialdehyde, a product of lipid peroxidation, is mutagenic in human cells. J Biol Chem 278: 31426-31433. doi: 10.1074/jbc.M212549200
|
| [29] |
Marnett LJ (2002) Oxy radicals, lipid peroxidation and DNA damage. Toxicology 181-182: 219-222. doi: 10.1016/S0300-483X(02)00448-1
|
| [30] |
Kadlubar FF, Anderson KE, Haussermann S, et al. (1998) Comparison of DNA adduct levels associated with oxidative stress in human pancreas. Mutat Res 405: 125-133. doi: 10.1016/S0027-5107(98)00129-8
|
| [31] |
Marnett LJ (1999) Lipid peroxidation-DNA damage by malondialdehyde. Mutat Res 424: 83-95. doi: 10.1016/S0027-5107(99)00010-X
|
| [32] |
Fink SP, Reddy GR, Marnett LJ (1997) Mutagenicity in Escherichia coli of the major DNA adduct derived from the endogenous mutagen malondialdehyde. Proc Natl Acad Sci USA 94: 8652-8657. doi: 10.1073/pnas.94.16.8652
|
| [33] |
Mao H, Schnetz-Boutaud NC, Weisenseel JP, et al. (1999) Duplex DNA catalyzes the chemical rearrangement of a malondialdehyde deoxyguanosine adduct. Proc Natl Acad Sci USA 96: 6615-6620. doi: 10.1073/pnas.96.12.6615
|
| [34] |
Mao H, Reddy GR, Manett LJ, et al. (1999) Solution structure of an oligodeoxynucleotide containing the malondialdehyde deoxyguanosine adduct N2-(3-oxo-1-propenyl)-dG (ring-opened M1G) positioned in a (CpG)3 frameshift hotspot of the Salmonella typhimurium hisD3052 gene. Biochemistry 38: 13491-13501. doi: 10.1021/bi9910124
|
| [35] |
Stone MP, Huang H, Brown KL, et al. (2011) Chemistry and structural biology of DNA damage and biological consequences. Chem Biodivers 8: 1571-1615. doi: 10.1002/cbdv.201100033
|
| [36] | Cohen SM, Garland EM, St John M, et al. (1992) Acrolein initiates rat urinary bladder carcinogenesis. Cancer Res 52: 3577-3581. |
| [37] |
Feng Z, Hu W, Hu Y, et al. (2006) Acrolein is a major cigarette-related lung cancer agent: preferential binding at p53 mutational hotspots and inhibition of DNA repair. Proc Natl Acad Sci USA 103: 15404-15409. doi: 10.1073/pnas.0607031103
|
| [38] |
Chung FL, Chen HJ, Nath RG (1996) Lipid peroxidation as a potential endogenous source for the formation of exocyclic DNA adducts. Carcinogenesis 17: 2105-2111. doi: 10.1093/carcin/17.10.2105
|
| [39] |
Chung FL, Nath RG, Nagao M, et al. (1999) Endogenous formation and significance of 1,N2-propanodeoxyguanosine adducts. Mutat Res 424: 71-81. doi: 10.1016/S0027-5107(99)00009-3
|
| [40] |
Nair U, Bartsch H, Nair J (2007) Lipid peroxidation-induced DNA damage in cancer-prone inflammatory diseases: a review of published adduct types and levels in humans. Free Radic Biol Med 43: 1109-1120. doi: 10.1016/j.freeradbiomed.2007.07.012
|
| [41] |
Zhang S, Villalta PW, Wang M, et al. (2007) Detection and quantitation of acrolein-derived 1,N2-propanodeoxyguanosine adducts in human lung by liquid chromatography-electrospray ionization-tandem mass spectrometry. Chem Res Toxicol 20: 565-571. doi: 10.1021/tx700023z
|
| [42] |
Yang IY, Hossain M, Miller H, et al. (2001) Responses to the major acrolein-derived deoxyguanosine adduct in Escherichia coli. J Biol Chem 276: 9071-9076. doi: 10.1074/jbc.M008918200
|
| [43] |
Yang IY, Johnson F, Grollman AP, et al. (2002) Genotoxic mechanism for the major acrolein-derived deoxyguanosine adduct in human cells. Chem Res Toxicol 15: 160-164. doi: 10.1021/tx010123c
|
| [44] |
de los Santos C, Zaliznyak T, Johnson F (2001) NMR characterization of a DNA duplex containing the major acrolein-derived deoxyguanosine adduct gamma -OH-1,-N2-propano-2'-deoxyguanosine. J Biol Chem 276: 9077-9082. doi: 10.1074/jbc.M009028200
|
| [45] |
Nair DT, Johnson RE, Prakash L, et al. (2008) Protein-template-directed synthesis across an acrolein-derived DNA adduct by yeast Rev1 DNA polymerase. Structure 16: 239-245. doi: 10.1016/j.str.2007.12.009
|
| [46] |
Zaliznyak T, Boonala R, Attaluri S, et al. (2009) Solution structure of DNA containing alpha-OH-PdG: the mutagenic adduct produced by acrolein. Nucleic Acid Res 37: 2153-2163. doi: 10.1093/nar/gkp076
|
| [47] |
Sanchez AM, Minko IG, Kurtz AJ, et al. (2003) Comparative evaluation of the bioreactivity and mutagenic spectra of acrolein-derived alpha-HOPdG and gamma-HOPdG regioisomeric deoxyguanosine adducts. Chem Res Toxicol 16: 1019-1028. doi: 10.1021/tx034066u
|
| [48] | Chung FL, Nath RG, Ocando J, et al. (2000) Deoxyguanosine adducts of t-4-hydroxy-2-nonenal are endogenous DNA lesions in rodents and humans: detection and potential sources. Cancer Res 60: 1507-1511. |
| [49] |
Cho YJ, Wang H, Kozekov ID, et al. (2006) Orientation of the crotonaldehyde-derived N2-[3-oxo-1(S)-methyl-propyl]-dG DNA adduct hinders interstrand cross-link formation in the 5'-CpG-3' sequence. Chem Res Toxicol 19: 1019-1029. doi: 10.1021/tx0600604
|
| [50] |
Brambilla G, Sciaba L, Faggin P, et al. (1986) Cytotoxicity, DNA fragmentation and sister-chromatid exchange in Chinese hamster ovary cells exposed to the lipid peroxidation product 4-hydroxynonenal and homologousaldehydes. Mutat Res 171: 169-176. doi: 10.1016/0165-1218(86)90051-0
|
| [51] |
Cajelli E, Ferraris A, Brambilla G (1987) Mutagenicity of 4-hydroxynonenal in V79 Chinese hamster cells. Mutat Res 190: 169-176. doi: 10.1016/0165-7992(87)90050-9
|
| [52] |
Eckl PM, Ortner A, Esterbauer H (1993) Genotoxic properties of 4-hydroxyalkenals and analogous aldehydes. Mutat res 290: 183-192. doi: 10.1016/0027-5107(93)90158-C
|
| [53] | Eckl P, Esterbauer H (1989) Genotoxic effects of 4-hydroxyalkenals. Adv Biosci 76: 141-157. |
| [54] |
Karlhuber GM, Bauer HC, Eckl PM (1997) Cytotoxic and genotoxic effects of 4-hydroxynonenal in cerebral endothelial cells. Mutat Res 381: 209-216. doi: 10.1016/S0027-5107(97)00170-X
|
| [55] |
Wacker M, Wanek P, Eder E (2001) Detection of q,N2-propano-deoxyguanosine adducts of trans-4-hydroxy-2-nonenal after gavage of trans-4-hydroxy-2-nonenal or induction of lipid peroxidation with carbon tetrachloride in F344 rats. Chem Biol Interact 137: 269-283. doi: 10.1016/S0009-2797(01)00259-9
|
| [56] |
Douki T, Odin F, Caillat S, et al. (2004) Predominance of the 1,N2-propano 2'-deoxyguanosine adduct among 4-hydroxy-2-nonenal-induced DNA lesions. Free Radic Biol Med 37: 62-70. doi: 10.1016/j.freeradbiomed.2004.04.013
|
| [57] |
Hu W, Feng Z, Eveleigh J, et al. (2002) The major lipid peroxidation product, trans-4-hydroxy-2-nonenal, preferentially forms DNA adducts at codon 249 of human p53 gene, a unique mutational hotspot in hepatocellular carcinoma. Carcinogenesis 23: 1781-1789. doi: 10.1093/carcin/23.11.1781
|
| [58] |
Huang H, Wang H, Qi N, et al. (2008) Rearrangement of the (6S,8R,11S) and (6R,8S,11R) exocyclic 1,N2-deoxyguanosine adducts of trans-4-hydroxynonenal to N2-deoxyguanosine cyclic hemiacetal adducts when placed complementary to cytosine in duplex DNA. J Am Chem Soc 130: 10898-10906. doi: 10.1021/ja801824b
|
| [59] |
Huang H, Wang H, Stephen Lloyd R, et al. (2009) Conformational interconversion of the trans-4-hydroxynonenal-derved (6S,8R,11S) 1,N2-deoxyguanosine adduct when mismatched with deoxyadenosine in DNA. Chem Res Toxicol 22: 187-200. doi: 10.1021/tx800320m
|
| [60] |
Kozekov ID, Nechev LV, Sanchez A, et al. (2001) Interchain cross-linking of DNA mediated by the principal adduct of acrolein. Chem Res Toxicol 14: 1482-1485. doi: 10.1021/tx010127h
|
| [61] |
Kozekov ID, Nechev LV, Moseley MS, et al. (2003) DNA interchain cross-links formed by acrolein and crotonaldehyde. J Am Chem Soc 125: 50-61. doi: 10.1021/ja020778f
|
| [62] |
Wang H, Kozekov ID, Harris TM, et al. (2003) Site-specific synthesis and reactivity of oligonucleotides containing stereochemically defined 1,N2-deoxyguanosine adducts of the lipid peroxidation product trans-4-hydroxynonenal. J Am Chem Soc 125: 5687-5700. doi: 10.1021/ja0288800
|
| [63] |
Cho YJ, Kim HY, Huang H, et al. (2005) Spectroscopic characterization of interstrand carbinolamine cross-links formed in the 5'-CpG-3' sequence by the acrolein-derived gamma-OH-1,N2-propano-2'-deoxyguanosine DNA adduct. J Am Chem Soc 127: 17686-17696. doi: 10.1021/ja053897e
|
| [64] |
Cho YJ, Wang H, Kozekov ID, et al. (2006) Stereospecific formation of interstrand carbinolamine DNA crosslinks by crotonaldehyde- and acetaldehyde-derived a-CH3-g-OH-1,N2-propano-2'-deoxyguanosine adducts in the 5'-CpG-3' sequence. Chem Res Toxicol 19: 195-208. doi: 10.1021/tx050239z
|
| [65] |
Chen HJC, Chung FL (1996) Epoxidation of trans-4-hydroxy-2-nonenal by fatty acid hydroperoxides and hydrogen peroxide. Chem Res Toxicol 9: 306-312. doi: 10.1021/tx9501389
|
| [66] | Sodum RS, Chung FL (1991) Stereoselective formation of in vitro nucleic acid adducts by 2,3-epoxy-4-hydroxynonanal. Cancer Res 51: 137-143. |
| [67] |
Chen HJ, Gonzalez FJ, Shou M, et al. (1998) 2,3-epoxy-4-hydroxynonanal, a potential lipid peroxidation product for etheno adduct formation, is not a substrate of human epoxide hydrolase. Carcinogenesis 19: 939-943. doi: 10.1093/carcin/19.5.939
|
| [68] |
Wei X, Yin H (2015) Covalent modification of DNA by α, β-unsaturated aldehydes derived from lipid peroxidation: Recent progress and challenges. Free Radic Res 49: 905-917. doi: 10.3109/10715762.2015.1040009
|
| [69] |
Kowalczyk P, Cieśla JM, Komisarski M, et al. (2004) Long-chain adducts of trans-4-hydroxy-2-nonenal to DNA bases cause recombination, base substitutions and frameshift mutations in M13 phage. Mutat Res 550: 33-48. doi: 10.1016/j.mrfmmm.2004.01.007
|
| [70] | Huang H, Kozekov ID, Kozekova A, et al. (2010) DNA cross-link induced by trans-4-hydroxynonenal. Environ Mol Mutagen 51: 625-634. |
| [71] |
Hussain SP, Raja K, Amstad PA, et al. (2000) Increased p53 mutation load in nontumorous human liver of wilson disease and hemochromatosis: oxyradical overload diseases. Proc Natl Acad Sci USA 97: 12770-12775. doi: 10.1073/pnas.220416097
|
| [72] |
Chung FL, Pan J, Choudhury S, et al. (2003) Formation of trans-4-hydroxy-2-nonenal- and other enal-derived cyclic DNA adducts from ω-3 and ω-6 polyunsaturated fatty acids and their roles in DNA repair and human p53 gene mutation. Mutat Res 531: 25-36. doi: 10.1016/j.mrfmmm.2003.07.001
|
| [73] |
Winczura A, Czubaty A, Winczura K, et al. (2014) Lipid peroxidation product 4-hydroxy-2-nonenal modulates base excision repair in human cells. DNA Repair (Amst) 22: 1-11. doi: 10.1016/j.dnarep.2014.06.002
|
| [74] |
Tang MS, Wang HT, Hu Y, et al. (2011) Acrolein induced DNA damage, mutagenicity and effect on DNA repair. Mol Nutr Food Res 55: 1291-1300. doi: 10.1002/mnfr.201100148
|
| [75] |
Munnia A, Amasio ME, Peluso M (2004) Exocyclic malondialdehyde and aromatic DNA adducts in larynx tissues. Free Radic Biol Med 37: 850-858. doi: 10.1016/j.freeradbiomed.2004.05.024
|
| [76] |
Peluso M, Munnia A, Risso GG, et al. (2011) Breast fine-needle aspiration malondialdehyde deoxyguanosine adduct in breast cancer. Free Radic Res 45: 477-482. doi: 10.3109/10715762.2010.549485
|
| [77] |
Lee SH, Williams MV, DuBois RN, et al. (2005) Cyclooxygenase-2-mediated DNA damage. J Biol Chem 280: 28337-28346. doi: 10.1074/jbc.M504178200
|
| [78] |
Matsuda T, Tao H, Goto M, et al. (2013) Lipid peroxidation-induced DNA adducts in human gastric mucosa. Carcinogenesis 34: 121-127. doi: 10.1093/carcin/bgs327
|
| [79] |
Lee HW, Wang HT, Weng MW, et al. (2014) Acrolein- and 4-Aminobiphenyl-DNA adducts in human bladder mucosa and tumor tissue and their mutagenicity in human urothelial cells. Oncotarget 5: 3526-3540. doi: 10.18632/oncotarget.1954
|
| [80] |
Parola M, Robino G, Marra F, et al. (1998) HNE interacts directly with JNK isoforms in human hepatic stellate cells. J Clin Invest 102: 1942-1950. doi: 10.1172/JCI1413
|
| [81] |
Ranjan D, Chen C, Johnston TD, et al. (2006) Stimulation of Epstein-Barr virus-infected human B cell growth by physiological concentrations of 4-hydroxynonenal. Cell Biochem Funct 24: 147-152. doi: 10.1002/cbf.1201
|
| [82] | Guéraud F (2017) 4-Hydroxynonenal metabolites and adducts in pre-carcinogenic conditions and cancer. Free Radic Biol Med pii: S0891-5849(16)31121-2. |
| [83] | Wu KC, Cui JY, Klaassen CD (2012) Effect of graded Nrf2 activation on phase-I and -II drug metabolizing enzymes and transporters in mouse liver. PLoS One: e39006. |
| [84] |
Morry J, Ngamcherdtrakul W, Yantasee W (2017) Oxidative stress in cancer and fibrosis: Opportunity for therapeutic intervention with antioxidant compounds, enzymes, and nanoparticles. Redox Biol 11: 240-253. doi: 10.1016/j.redox.2016.12.011
|
| [85] | Poljšak B, Fink R (2014) The protective role of antioxidants in the defence against ROS/RNS-mediated environmental pollution. Oxid Med Cell Longev 2014: 671539. |
| [86] |
Pelicano H, Carney D, Huang P (2004) ROS stress in cancer cells and therapeutic implications. Drug Resist 7: 97-110. doi: 10.1016/j.drup.2004.01.004
|
| [87] | Young O, Crotty T, O'Connell R, et al. (2010) Levels of oxidative damage and lipid peroxidation inthyroidneoplasia. Head and Neck 32: 750-756. |
| [88] |
Patel BP, Rawal UM, Dave TK, et al. (2007) Lipid peroxidation, total antioxidant status, and total thiol levels predict overall survival in patients with oral squamous cell carcinoma. Integrative Cancer Therapies 6: 365-372. doi: 10.1177/1534735407309760
|
| [89] |
Tsao SM, Yin MC, Liu WH (2007) Oxidant stress and B vitamins status in patients with non-small cell lung cancer. Nutr Cancer 59: 8-13. doi: 10.1080/01635580701365043
|
| [90] |
Kumar B, Koul S, Khandrika L, et al. (2008) Oxidative stress is inherent in prostate cancer cells and is required for aggressive phenotype. Cancer Res 68: 1777-1785. doi: 10.1158/0008-5472.CAN-07-5259
|
| [91] |
Fruehauf JP, Meyskens FL Jr. (2007) Reactive oxygen species: a breath of life or death? Clin Cancer Res 13: 789-794. doi: 10.1158/1078-0432.CCR-06-2082
|
| [92] | Barrera G (2012) Oxidative Stress and Lipid Peroxidation Products in Cancer Progression and Therapy. ISRN Oncology: 137289. |
| [93] |
Forman HJ, Fukuto JM, Miller T, et al. (2008) The chemistry of cell signaling by reactive oxygen and nitrogen species and 4-hydroxynonenal. Arch Biochem Biophys 477: 183-195. doi: 10.1016/j.abb.2008.06.011
|
| [94] | Lushchak VI (2012) Glutathione homeostasis and functions: potential targets for medical interventions. J Amino Acid, 2012. |
| [95] |
Awasthi SS, Singhal YC, Awasthi B, et al. (2008) RLIP76 and cancer. Clin Cancer Res 14: 4372-4377. doi: 10.1158/1078-0432.CCR-08-0145
|
| [96] |
Tjalkens RB, Cook LW, Petersen DR (1999) Formation and export of the glutathione conjugate of 4-hydroxy-2,3-Enonenal(4-HNE) in hepatoma cells. Arch Biochem Biophys 361: 113-119. doi: 10.1006/abbi.1998.0946
|
| [97] | Gasparovic AC, Milkovic L, Sunjic SB, et al. (2017) Cancer growth regulation by 4-hydroxynonenal. Free Radic Biol Med pii: S0891-5849(17)30039-4. |
| [98] |
Zarkovic N, Ilic Z, Jurin M, et al. (1993) Stimulation of HeLa cell growth by physiological concentrations of 4-hydroxynonenal. Cell Biochem. Funct 11: 279-286. doi: 10.1002/cbf.290110409
|
| [99] |
Zanetti D, Poli G, Vizio B, et al. (2003) 4-hydroxynonenal and transforming growth factor-beta1 expression in colon cancer. Mol Aspects Med 24: 273-280. doi: 10.1016/S0098-2997(03)00022-0
|
| [100] |
Cerbone A, Toaldo C, Laurora S, et al. (2007) 4-Hydroxynonenal and PPARgamma ligands affect proliferation, differentiation, and apoptosis in colon cancer cells. Free Radic Biol Med 42: 1661-1670. doi: 10.1016/j.freeradbiomed.2007.02.009
|
| [101] |
Barrera G, Muraca R, Pizzimenti S, et al. (1994) Inhibition of c-myc expression induced by 4-hydroxynonenal, a product of lipid peroxidation, in the HL-60 human leukemic cell line. Biochem Biophys Res Commun 203: 553-561. doi: 10.1006/bbrc.1994.2218
|
| [102] |
Pizzimenti S, Barrera G, Dianzani MU, et al. (1999) Inhibition of D1, D2, and A-cyclin expression in HL-60 cells by the lipid peroxydation product 4-hydroxynonenal. Free Radic Biol Med 26: 1578-1586. doi: 10.1016/S0891-5849(99)00022-2
|
| [103] |
Pizzimenti S, Briatore F, Laurora S, et al. (2006) 4-Hydroxynonenal inhibits telomerase activity and hTERT expression in human leukemic cell lines. Free Radic Biol Med 40: 1578-1591. doi: 10.1016/j.freeradbiomed.2005.12.024
|
| [104] |
Albright CD, Klem E, Shah AA, et al. (2005) Breast cancer cell-targeted oxidative stress: enhancement of cancer cell uptake of conjugated linoleic acid, activation of p53, and inhibition of proliferation. Exp Mol Pathol 79: 118-125. doi: 10.1016/j.yexmp.2005.05.005
|
| [105] |
Sunjic SB, Cipak A, Rabuzin F, et al. (2005) The influence of 4-hydroxy-2-nonenal on proliferation, differentiation and apoptosis of human osteosarcoma cells. Biofactors 24: 141-148. doi: 10.1002/biof.5520240117
|
| [106] |
Pettazzoni P, Pizzimenti S, Toaldo C, et al. (2011) Induction of cell cycle arrest and DNA damage by the HDAC inhibitor panobinostat (LBH589) and the lipid peroxidation end product 4-hydroxynonenal in prostate cancer cells. Free Radic Biol Med 50: 313-322. doi: 10.1016/j.freeradbiomed.2010.11.011
|
| [107] |
Chaudhary P, Sharma R, Sahu M, et al. (2013) 4-Hydroxynonenal induces G2/M phase cell cycle arrest by activation of the ataxia telangiectasia mutated and Rad3-related protein (ATR)/checkpoint kinase 1 (Chk1) signaling pathway. J Biol Chem 288: 20532-20546. doi: 10.1074/jbc.M113.467662
|
| [108] | Ji GR, Yu NC, Xue X, et al. (2014) 4-Hydroxy-2-nonenal induces apoptosis by inhibiting AKT signaling in human osteosarcoma cells. Scientific World Journal 2014: 873525. |
| [109] | Cao ZG, Xu X, Xue YM, et al. (2014) Comparison of 4-hydroxynonenal-induced p53-mediated apoptosis in prostate cancer cells LNCaP and DU145. Contemp Oncol (Pozn) 18: 22-28. |
| [110] |
Bauer G, Zarkovic N (2015) Revealing mechanisms of selective, concentration-dependent potentials of 4-hydroxy-2-nonenal to induce apoptosis in cancer cells through inactivation of membrane-associated catalase. Free Radic Biol Med 81: 128-144. doi: 10.1016/j.freeradbiomed.2015.01.010
|
| [111] |
Dasari S, Tchounwou PB (2014) Cisplatin in cancer therapy: molecular mechanisms of action. Eur J Pharmacol 740: 364-378. doi: 10.1016/j.ejphar.2014.07.025
|
| [112] |
Thorn CF, Oshiro C, Marsh S, et al. (2011) Doxorubicin pathways: pharmacodynamics and adverse effects. Pharmacogenet Genomics 21: 440-446. doi: 10.1097/FPC.0b013e32833ffb56
|
| [113] |
Casares C, Ramírez-Camacho R, Trinidad A, et al. (2012) Reactive oxygen species in apoptosis induced by cisplatin: review of physiopathological mechanisms in animal models. Eur Arch Otorhinolaryngol 269: 2455-2459. doi: 10.1007/s00405-012-2029-0
|
| [114] |
Cipak A, Jaganjac M, Tehlivets O, et al. (2008) Adaptation to oxidative stress induced by polyunsaturated fatty acids in yeast. Biochim Biophys Acta 1781: 283-287. doi: 10.1016/j.bbalip.2008.03.010
|
| [115] |
Awasthi YC, Yang Y, Tiwari NK, et al. (2004) Regulation of 4-hydroxynonenal-mediated signaling by glutathione S-transferases. Free Radic Biol Med 37: 607-619. doi: 10.1016/j.freeradbiomed.2004.05.033
|
| [116] | Traverso N, Ricciarelli R, Nitti M, et al. (2013) Role of glutathione in cancer progression and chemoresistance. Oxid Med Cell Longev 2013: 972913. |
| [117] | Furfaro AL, Traverso N, Domenicotti C, et al. (2016) The Nrf2/HO-1 axis in cancer cell growth and chemoresistance. Oxid Med Cell Longev 2016: 1958174. |
| [118] |
Trachootham D, Alexandre J, Huang P (2009) Targeting cancer cells by ROS-mediated mechanisms: a radical therapeutic approach? Nat Rev Drug Discov 8: 579-591. doi: 10.1038/nrd2803
|
| [119] |
Trachootham D, Zhou Y, Zhang H, et al. (2006) Selective killing of oncogenically transformed cells through a ROS-mediated mechanism by beta-phenylethyl isothiocyanate. Cancer Cell 10: 241-252. doi: 10.1016/j.ccr.2006.08.009
|
| [120] |
Trachootham D, Zhang H, Zhang W, et al. (2008) Effective elimination of fludarabine-resistant CLL cells by PEITC through a redox-mediated mechanism. Blood 112: 1912-1922. doi: 10.1182/blood-2008-04-149815
|
| [121] |
Zhang H, Trachootham D, Lu W, et al. (2008) Effective killing of Gleevec-resistant CML cells with T315I mutation by a natural compound PEITC through redox-mediated mechanism. Leukemia 22: 1191-1199. doi: 10.1038/leu.2008.74
|
| [122] |
Oberley TD, Toyokuni S, Szweda LI (1999) Localization of hydroxyl-nonenal protein adducts in normal human kidney and selected human kidney cancers. Free Radic Biol Med 27: 695-703. doi: 10.1016/S0891-5849(99)00117-3
|
| [123] |
Skrzydlewska E, Stankiewicz A, Sulkowska M, et al (2001) Antioxidant status and lipid peroxidation in colorectal cancer. J Toxicol Environ Health 64: 213-222. doi: 10.1080/15287390152543690
|
| [124] | Juric-Sekhar G, Zarkovic K, Waeg G, et al. (2009) Distribution of 4-hydroxynonenal-protein conjugates as a marker of LPO and parameter of malignancy in astrocytic and ependymal tumors of the brain. Tumori 95: 762-768. |
| [125] |
Karihtala P, Kauppila S, Puistola U, et al. (2011) Divergent behaviour of oxidative stress markers 8-hydroxydeoxyguanosine (8-OHdG) and 4-hydroxy-2 nonenal (HNE) in breast carcinogenesi. Histopathology 58: 854-862. doi: 10.1111/j.1365-2559.2011.03835.x
|
| [126] |
Dianzani MU (2003) 4-Hydroxynonenal from pathology to physiology. Mol Aspects Med 24: 263-272. doi: 10.1016/S0098-2997(03)00021-9
|
| [127] |
Uchida K (2003) 4-Hydroxy-2-nonenal: a product and mediator of oxidative stress. Prog Lipid Res 42: 318-343. doi: 10.1016/S0163-7827(03)00014-6
|
| [128] |
Cohen G, Riahi Y, Sasson S (2011) Lipid peroxidation of poly-unsaturated fatty acids in normal and obese adipose tissues. Arch Physiol Biochem 117: 131-139. doi: 10.3109/13813455.2011.557387
|
| [129] |
White MF (2002) IRS proteins and the common path to diabetes. Am J Physiol Endocrinol Metab 283: 413-422. doi: 10.1152/ajpendo.00514.2001
|
| [130] |
Leonarduzzi G, Chiarpotto E, Biasi F, et al. (2005) 4-Hydroxynonenal and cholesterol oxidation products in atherosclerosis. Mol Nutr Food Res 49: 1044-1049. doi: 10.1002/mnfr.200500090
|
| [131] | Arcaro A, Daga M, Cetrangolo GP, et al. (2015) Generation of adducts of 4-hydroxy-2-nonenal with heat shock 60 kDa protein 1 in human prolyelocytic HL-60 and monocytic THP-1 cell lines. Oxid Med Cell Longev 2015: 296146. |
| [132] | Calderwood SK, Stevenson MA, Murshid A (2012) Heat shock proteins, autoimmunity, and cancer treatment. Autoimmune Dis 2012: 486069 |
| [133] |
Grundtman C, Kreutmayer SB, Almanzar G, et al. (2011) Heat shock protein 60 and immune inflammatory responses in atherosclerosis. Arterioscler Thromb Vasc Biol 31: 960-968 doi: 10.1161/ATVBAHA.110.217877
|
| [134] | Nègre-Salvayre A, Garoby-Salom S, et al. (2016) Proatherogenic effects of 4-hydroxynonenal. Free Radic Biol Med pii: S0891-5849(16)31138-8. |
| [135] | Gargiulo S, Testa G, Gamba P, et al. (2017) Oxysterols and 4-hydroxy-2-nonenal contribute to atherosclerotic plaque destabilization. Free Radic Biol Med pii: S0891-5849(16)31139-X. |
| [136] | Li CJ, Nanji AA, Siakotos AN, et al. (1997) Acetaldehyde-modified and 4-hydroxynonenal-modified proteins in the livers of rats with alcoholic liver disease. Hepatology 26: 650-657. |
| [137] |
McKim SE, Uesugi T, Raleigh JA, et al. (2003) Chronic intragastric alcohol exposure causes hypoxia and oxidative stress in the rat pancreas. Arch Biochem Biophys 417: 34-43. doi: 10.1016/S0003-9861(03)00349-7
|
| [138] |
Turk PW, Laayoun A, Smith SS, et al. (1995) DNA adduct 8-hydroxyl-2′-deoxyguanosine (8-hydroxyguanine) affects function of human DNA methyltransferase. Carcinogenesis 16: 1253-1255. doi: 10.1093/carcin/16.5.1253
|
| [139] |
Nair J, Godschalk RW, Nair U, et al. (2012) Identification of 3,N(4)-etheno-5-methyl-2'-deoxycytidine in human DNA: a new modified nucleoside which may perturb genome methylation. Chem Res Toxicol 25: 162-169. doi: 10.1021/tx200392a
|
| [140] |
Yara S, Lavoie JC, Beaulieu JF, et al. (2013) Iron-ascorbate-mediated lipid peroxidation causes epigenetic changes in the antioxidant defense in intestinal epithelial cells: impact on inflammation. PLoS One 8: e63456. doi: 10.1371/journal.pone.0063456
|
| [141] |
Nair J, Gansauge F, Beger H, et al. (2006) Increased etheno-DNA adducts in affected tissues of patients suffering from Crohn's disease, ulcerative colitis, and chronic pancreatitis. Antioxid Redox Signal 8: 1003-1010. doi: 10.1089/ars.2006.8.1003
|
| [142] |
Nair J, Srivatanakul P, Haas C, et al. (2010) High urinary excretion of lopid peroxidation-derived DNA damage in patients with cancer-prone liver diseases. Mutat Res 683: 23-28. doi: 10.1016/j.mrfmmm.2009.10.002
|
| [143] | Pizzimenti S, Ciamporcero E, Daga M, et al. (2013) Interaction of aldehydes derived from lipid peroxidation and membrane proteins. Frontiers Physiol 4: 242. |
| [144] | Barrera G, Pizzimenti S, Daga M, et al. (2016) Aldehydes derived from lipid peroxidation in cancer and autoimmunity. In: Lipid Peroxidation: Inhibition, Effects and Mechanisms (Angel Catalá, Ed.), Nova Science Publishers, Inc., New York, 2017. |
| [145] |
Weismann D, Binder CJ (2012) The innate immune response to products of phospholipid peroxidation. Biochim Biophysica Acta 1818: 2465-2475. doi: 10.1016/j.bbamem.2012.01.018
|
| [146] |
Chou MY, Fogelstrand L, Hartvigsen K, et al. (2009) Oxidation-specific epitopes are dominant targets of innate natural antibodies in mice and humans. J Clin Invest 119: 1335-1349. doi: 10.1172/JCI36800
|
| [147] |
Ohki I, Ishigaki T, Oyama T, et al. (2005) Crystal structure of human lectin-like, oxidized low-density lipoprotein receptor 1 ligand binding domain and its ligand recognition mode to OxLDL. Structure 13: 905-917. doi: 10.1016/j.str.2005.03.016
|
| [148] |
Nickel T, Schmauss D, Hanssen H, et al. (2009) OxLDL uptake by dendritic cells induces upregulation of scavenger-receptors, maturation and differentiation. Atherosclerosis 205: 442-450. doi: 10.1016/j.atherosclerosis.2009.01.002
|
| [149] |
Wuttge DM, Bruzelius M, Stemme S (1999). T-cell recognition of lipid peroxidation products breaks tolerance to self proteins. Immunology 98: 273-279. doi: 10.1046/j.1365-2567.1999.00872.x
|
| [150] |
Scofield RH, Kurien BT, Ganick S, et al. (2005) Modification of lupus-associated 60-kDa Ro protein with the lipid oxidation product 4-hydroxy-2-nonenal increases antigenicity and facilitates epitope spreading. Free Radic Biol Med 38: 719-728. doi: 10.1016/j.freeradbiomed.2004.11.001
|
| [151] |
Kurien BT, Hensley K, Bachmann M, et al. (2006) Oxidatively modified autoantigens in autoimmune diseases. Free Radic Biol Med 41: 549-556. doi: 10.1016/j.freeradbiomed.2006.05.020
|
| [152] |
Kurien BT, Scofield RH (2006) Autoantibody determination in the diagnosis of systemic lupus erythematosus. Scand J Immunol 64: 227-235. doi: 10.1111/j.1365-3083.2006.01819.x
|
| [153] |
Casciola-Rosen LA, Anhalt G, Rosen A (1994) Autoantigens targeted in sistemic lupus erythematosus are clustered in two populations of surface structures on apoptotic keratinocytes. J Exp Med 179: 1317-1330. doi: 10.1084/jem.179.4.1317
|
| [154] |
Miranda-Carús M-E, Askanase AD, Clancy RM, et al. (2000) Anti-SSA/Ro and anti-SSB/La autoantibodies bind the surface of apoptotic fetal cardiocytes and promote secretion of TNF-a by macrophages. J Immunol 165: 5345-5351. doi: 10.4049/jimmunol.165.9.5345
|
| [155] |
Savill J, Dransfield I, Gregory C, et al. (2002) A blast from the past: clearance of apoptotic cells regulates immune responses. Nat Rev Immunol 2: 965-975. doi: 10.1038/nri957
|
| [156] | Emlen W, Niebur J, Kadera R (1994) Accelerated in vitro apoptosys of lymphocytes from patients with systemic lupus erythematosus. J Immunol 152: 3685-3692 |
| [157] |
Georgescu L, Vakkalanka RK, Elkon KB, et al. (1997) Interleukin-10 promotes activation-induced cell death of SLE lymphocytes mediated by Fas ligand. J Clin Invest 100: 2622-2633. doi: 10.1172/JCI119806
|
| [158] |
Ren Y, Tang J, Mok MY, et al. (2003) Increased apoptotic neutrophils and macrophages and impaired macrophage phagocytic clearance of apoptotic neutrophils in systemic lupus erythematosus. Arthritis Rheum 48: 2888-2897. doi: 10.1002/art.11237
|
| [159] |
Zeher M, Szodoray P, Gyimesi E, et al. (1999) Correlation of increased susceptibility to apoptosis of CD4+ T cells with lymphocyte activation and activity of disease in patients with primary Sjögren's syndrome. Arthritis Rheum 42: 1673-1681. doi: 10.1002/1529-0131(199908)42:8<1673::AID-ANR16>3.0.CO;2-1
|
| [160] |
Espinosa A, Zhou W, Ek M, et al. (2006) The Sjögren's Sydrome-associated autoantigen Ro52 is an E3 ligase that regulates proliferation and cell death. J Immunol 176: 6277-6285. doi: 10.4049/jimmunol.176.10.6277
|
| [161] |
Licht R, Dieker JWC, Jacobs CWM, et al. (2004) Decreased phagocytosis of apoptotic cells in diseased SLE mice. J Autoimmun 22: 139-145. doi: 10.1016/j.jaut.2003.11.003
|
| [162] |
Pan Z-J, Davis K, Maier S, et al. (2006) Neo-epitopes are required for immunogenicity of the La/SS-B nuclear antigen in the context of late apoptotic cells. Clin Experim Immunol 143: 237-248. doi: 10.1111/j.1365-2249.2005.03001.x
|
| [163] |
Kurien T, Porter A, Dorri Y, et al. (2011) Degree of modification of Ro60 by the lipid peroxidation by-product 4-hydroxy-2-nonenal may differentially induce Sjögren's syndrome or sistemic lupus erythematosus in BALB/c mice. Free Radic Biol Med 50: 1222-1233. doi: 10.1016/j.freeradbiomed.2010.10.687
|
| [164] |
Khan F, Moinuddin, Mir AR, et al. (2016) Immunochemical studies on HNE-modified HSA: Anti-HNE-HSA antibodies as a probe for HNE damaged albumin in SLE. Int J Biol Macromol 86: 145-154. doi: 10.1016/j.ijbiomac.2016.01.053
|
| [165] |
Khatoon F, Moinuddin, Alam K, et al. (2012) Physicochemical and immunological studies on 4-hydroxynonenal modified HSA: Implications of protein damage by lipid peroxidation products in the etiopathogenesis of SLE. Hum Immunol 73: 1132-1139. doi: 10.1016/j.humimm.2012.08.011
|
| [166] | Chen Q, Esterbauer H, Jurgens H (1992) Studies on epitopes on low-density lipoprotein modified by 4-hydroxynonenal. Biochemical characterization and determination. Biochemical J 288: 249-254. |
| [167] |
Uchida K, Szweda LI, Chae HZ, et al. (1993) Immunochemical detection of 4-hydroxynonenal protein adducts in oxidized hepatocytes. Proc Natl Acad Sci USA 90: 8742-8746. doi: 10.1073/pnas.90.18.8742
|
| [168] | Hashimoto M, Shibata T, Wasada H, et al. (2003) Structural basis of protein-bound endogenous aldehydes. Chemical and immunochemical characterization of configurational isomers of a 4-hydroxy-2-nonenal-histidine adduct. J Biol Chem 278: 5044-5051. |
| [169] |
Akagawa M, Ito S, Toyoda K, et al. (2006) Bispecific Abs against modified protein and DNA with oxidized lipids. Proc Natl Acad Sci USA 103: 6160-6165. doi: 10.1073/pnas.0600865103
|
| [170] | Toyoda K, Nagae R, Akagawa M, et al. (2007) Protein-bound 4-hydroxy-2-nonenal. An endogenous triggering antigen of anti-DNA response. J Biol Chem 282: 25769-25778. |
| [171] |
Al-Shobaili HA, Al Robaee AA, Alzolibani AA, et al. (2012) Antibodies against 4-hydroxy-2-nonenal modified epitopes recognized chromatin and its oxidized forms: role of chromatin, oxidized forms of chromatin and 4-hydroxy-2-nonenal modified epitopes on the etiopathogenesis of SLE. Disease Markers 33: 19-34. doi: 10.1155/2012/532497
|
| [172] |
Seki S, Kitada T, Yamada T, et al. (2002) In situ detection of lipid peroxidation and oxidative DNA damage in non-alcoholic fatty liver disease. J Hepatol 37: 56-62. doi: 10.1016/S0168-8278(02)00073-9
|
| [173] |
Chalasani N, Deeg MA, Crabb DW (2004) Systemic lipid peroxidation and its metabolic and dietary correlates in patients with non-alcoholic steatohepatitis. Am J Gastroenterol 99: 1497-1502. doi: 10.1111/j.1572-0241.2004.30159.x
|
| [174] | Nobili V, Parola M, Alisi A, et al. (2010) Oxidative stress parameters in paediatric non-alcoholic fatty liver disease. Int J Molec Med 26: 471-476. |
| [175] | Teufel U, Peccerella T, Engelmann G, et al. (2015) Detection of carcinogenic etheno-DNA adducts in children and adolescents with non-alcoholic steatohepatitis (NASH). Hepatobiliary Surg Nutr 4: 426-35. |
| [176] | Ogawa M, Matsuda T, Ogata A, et al. (2013) DNA damage in rheumatoid arthritis: an age-dependent increase in the lipid peroxidation-derived DNA adduct, heptanone-etheno-2'-deoxycytidine. Autoimmune Dis 2013: 183487. |
| [177] | Wang G, Pierangeli SS, Papalardo E, et al. (2010) Markers of oxidative and nitrosative stress in systemic lupus erythematosus: correlation with disease activity. Arthritis Rheum 62: 2064-2072. |

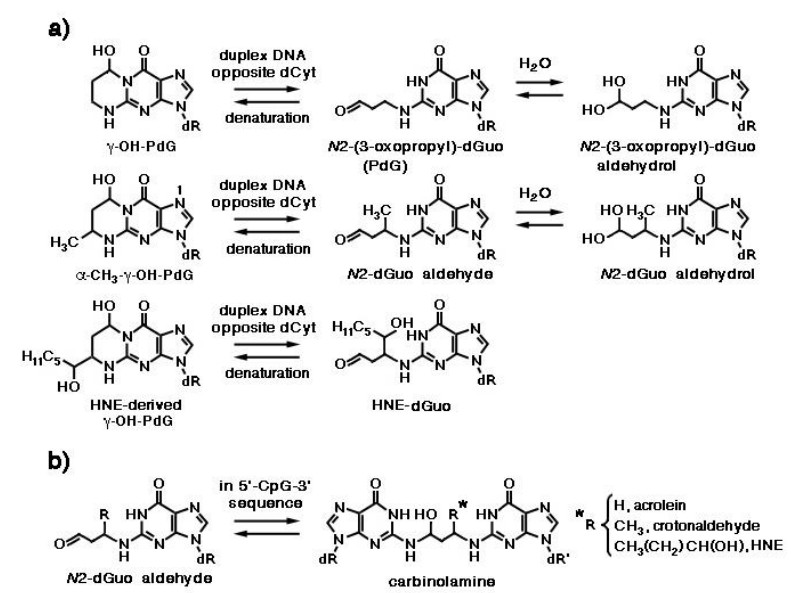

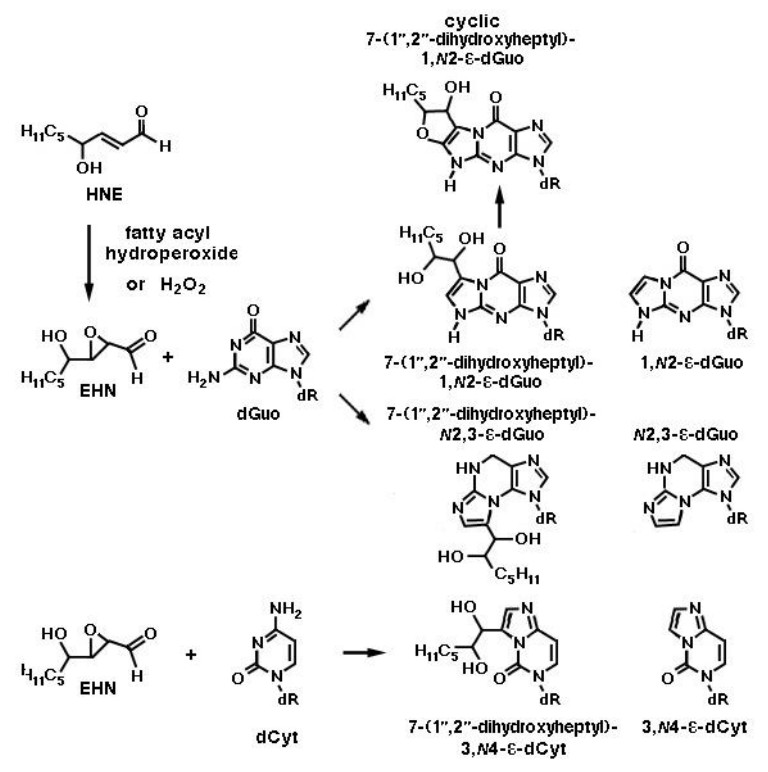
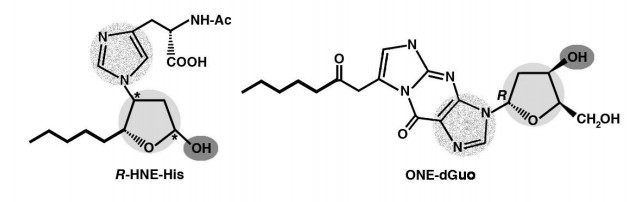
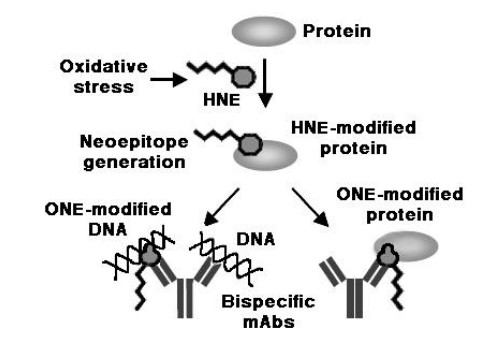
Figures(10) / Tables(1)

Fabrizio Gentile, Alessia Arcaro, Stefania Pizzimenti, Martina Daga, Giovanni Paolo Cetrangolo, Chiara Dianzani, Alessio Lepore, Maria Graf, Paul R. J. Ames, Giuseppina Barrera. DNA damage by lipid peroxidation products: implications in cancer, inflammation and autoimmunity[J]. AIMS Genetics, 2017, 4(2): 103-137. doi: 10.3934/genet.2017.2.103







 DownLoad:
DownLoad: