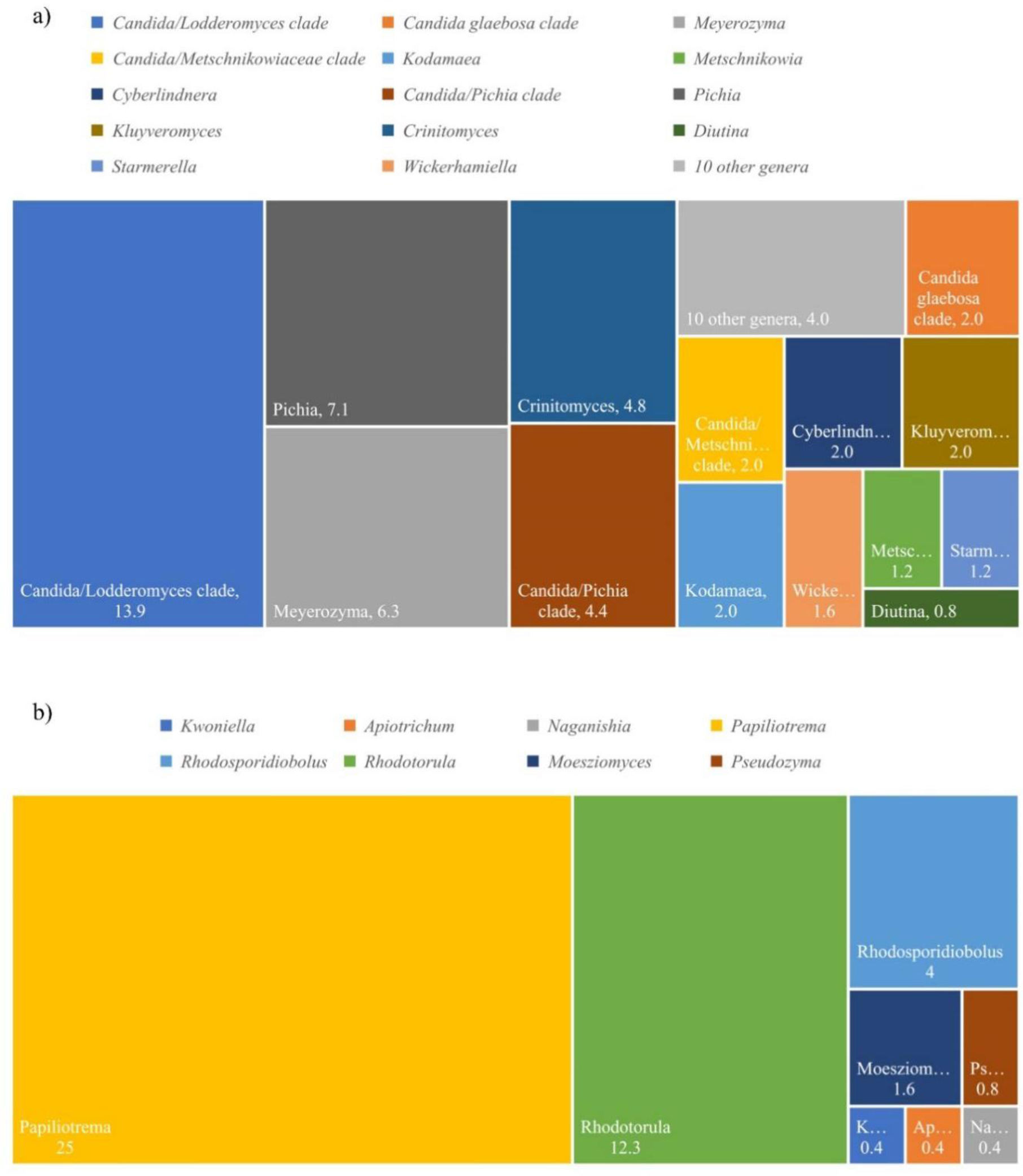
The diversity of duckweed (Lemnaceae) associated yeasts was studied using a culture-dependent method. A total of 252 yeast strains were isolated from 53 duckweed samples out of the 72 samples collected from 16 provinces in Thailand. Yeast identification was conducted based on the D1/D2 region of the large subunit (LSU) rRNA gene sequence analysis. It revealed that 55.2% and 44.8% yeast species were Ascomycota and Basidiomycota duckweed associated yeasts, respectively. Among all, Papiliotrema laurentii, a basidiomycetous yeast, was found as the most prevalent species showing a relative of frequency and frequency of occurrence of 21.8% and 25%, respectively. In this study, high diversity index values were shown, indicated by the Shannon-Wiener index (H′), Shannon equitability index (EH) and Simpson diversity index (1-D) values of 3.48, 0.86 and 0.96, respectively. The present results revealed that the yeast community on duckweed had increased species diversity, with evenness among species. Principal coordinate analysis (PCoA) revealed no marked differences in yeast communities among duckweed genera. The species accumulation curve showed that the observed species richness was lower than expected. Investigation of the plant growth promoting traits of the isolated yeast on duckweed revealed that 178 yeast strains produced indole-3-acetic acid (IAA) at levels ranging from 0.08–688.93 mg/L. Moreover, siderophore production and phosphate solubilization were also studied. One hundred and seventy-three yeast strains produced siderophores and exhibited siderophores that showed 0.94–2.55 activity units (AU). One hundred six yeast strains showed phosphate solubilization activity, expressed as solubilization efficiency (SE) units, in the range of 0.32–2.13 SE. This work indicates that duckweed associated yeast is a potential microbial resource that can be used for plant growth promotion.
Citation: Napapohn Kajadpai, Jirameth Angchuan, Pannida Khunnamwong, Nantana Srisuk. Diversity of duckweed (Lemnaceae) associated yeasts and their plant growth promoting characteristics[J]. AIMS Microbiology, 2023, 9(3): 486-517. doi: 10.3934/microbiol.2023026
The diversity of duckweed (Lemnaceae) associated yeasts was studied using a culture-dependent method. A total of 252 yeast strains were isolated from 53 duckweed samples out of the 72 samples collected from 16 provinces in Thailand. Yeast identification was conducted based on the D1/D2 region of the large subunit (LSU) rRNA gene sequence analysis. It revealed that 55.2% and 44.8% yeast species were Ascomycota and Basidiomycota duckweed associated yeasts, respectively. Among all, Papiliotrema laurentii, a basidiomycetous yeast, was found as the most prevalent species showing a relative of frequency and frequency of occurrence of 21.8% and 25%, respectively. In this study, high diversity index values were shown, indicated by the Shannon-Wiener index (H′), Shannon equitability index (EH) and Simpson diversity index (1-D) values of 3.48, 0.86 and 0.96, respectively. The present results revealed that the yeast community on duckweed had increased species diversity, with evenness among species. Principal coordinate analysis (PCoA) revealed no marked differences in yeast communities among duckweed genera. The species accumulation curve showed that the observed species richness was lower than expected. Investigation of the plant growth promoting traits of the isolated yeast on duckweed revealed that 178 yeast strains produced indole-3-acetic acid (IAA) at levels ranging from 0.08–688.93 mg/L. Moreover, siderophore production and phosphate solubilization were also studied. One hundred and seventy-three yeast strains produced siderophores and exhibited siderophores that showed 0.94–2.55 activity units (AU). One hundred six yeast strains showed phosphate solubilization activity, expressed as solubilization efficiency (SE) units, in the range of 0.32–2.13 SE. This work indicates that duckweed associated yeast is a potential microbial resource that can be used for plant growth promotion.

| [1] |
Wang W, Wu Y, Yan Y, et al. (2010) DNA barcoding of the Lemnaceae, a family of aquatic monocots. BMC Plant Biol 10: 1-11. https://doi.org/10.1186/1471-2229-10-205 
|
| [2] | Landolt E, Kandeler R (1987) Biosystematic investigations in the family of duckweeds (Lemnaceae), Vol. 4: the family of Lemnaceae-a monographic study, Vol. 2 (phytochemistry, physiology, application, bibliography). Veroeffentlichungen des Geobotanischen Instituts der ETH, Stiftung Ruebel (Switzerland) . https://doi.org/10.1007/BF00037640 |
| [3] |
Tippery NP, Les DH (2020) Tiny plants with enormous potential: phylogeny and evolution of Duckweeds. The Duckweed Genomes . Cham: Springer International Publishing 19-38. https://doi.org/10.1007/978-3-030-11045-1_2
|
| [4] |
Sońta M, Rekiel A, Batorska M (2019) Use of duckweed (Lemna L.) in sustainable livestock production and aquaculture–a review. Ann Anim Sci 19: 257-271. https://doi.org/10.2478/aoas-2018-0048
|
| [5] |
Bhanthumnavin K, Mcgarry MG (1971) Wolffia arrhiza as a possible source of inexpensive protein. Nature 232: 495-495. https://doi.org/10.1038/232495a0
|
| [6] |
Appenroth KJ, Sree KS, Böhm V, et al. (2017) Nutritional value of duckweeds (Lemnaceae) as human food. Food Chem 217: 266-273. https://doi.org/10.1016/j.foodchem.2016.08.116
|
| [7] |
Edelman M, Colt M (2016) Nutrient value of leaf vs. seed. Front Chem 4: 32. https://doi.org/10.3389/fchem.2016.00032
|
| [8] |
Appenroth KJ, Sree KS, Bog M, et al. (2018) Nutritional value of the duckweed species of the genus Wolffia (Lemnaceae) as human food. Front Chem 6: 483. https://doi.org/10.3389/fchem.2018.00483
|
| [9] |
Acosta K, Appenroth KJ, Borisjuk L, et al. (2021) Return of the Lemnaceae: Duckweed as a model plant system in the genomics and postgenomics era. Plant Cell 33: 3207-3234. https://doi.org/10.1093/plcell/koab189
|
| [10] | Yinglin W, Yuanyuan Y, Qian X, et al. (2022) Research advances on application of duckweed in bioremediation of polluted water. Ecotoxicol Environ Saf 74–85. https://doi.org/10.7524/AJE.1673-5897.20210831002 |
| [11] |
Cedergreen N, Madsen TV (2002) Nitrogen uptake by the floating macrophyte Lemna minor. New Phytol 155: 285-292. https://doi.org/10.1046/j.1469-8137.2002.00463.x
|
| [12] |
Yan A, Wang Y, Tan SN, et al. (2020) Phytoremediation: a promising approach for revegetation of heavy metal-polluted land. Front Plant Sci 11: 359. https://doi.org/10.3389/fpls.2020.00359
|
| [13] |
Ziegler P, Sree K, Appenroth KJ (2016) Duckweeds for water remediation and toxicity testing. Toxicol Environ Chem 98: 1127-1154. https://doi.org/10.1080/02772248.2015.1094701
|
| [14] |
Zhou H, Liu X, Chen X, et al. (2018) Characteristics of removal of waste-water marking pharmaceuticals with typical hydrophytes in the urban rivers. Sci Total Environ 636: 1291-1302. https://doi.org/10.1016/j.scitotenv.2018.04.384
|
| [15] |
Ekperusi AO, Nwachukwu EO, Sikoki FD (2020) Assessing and modelling the efficacy of Lemna paucicostata for the phytoremediation of petroleum hydrocarbons in crude oil-contaminated wetlands. Sci Rep 10: 1-9. https://doi.org/10.1038/s41598-020-65389-z
|
| [16] |
Osama R, Awad HM, Ibrahim MG, et al. (2020) Mechanistic and economic assessment of polyester wastewater treatment via baffled duckweed pond. J Water Process Eng 35: 101179. https://doi.org/10.1016/j.jwpe.2020.101179
|
| [17] |
Cheng JJ, Stomp AM (2009) Growing duckweed to recover nutrients from wastewaters and for production of fuel ethanol and animal feed. Clean-Soil Air Water 37: 17-26. https://doi.org/10.1002/clen.200800210
|
| [18] |
Cui W, Cheng J (2015) Growing duckweed for biofuel production: a review. Plant Biol 17: 16-23. https://doi.org/10.1111/plb.12216
|
| [19] |
Guo L, Jin Y, Xiao Y, et al. (2020) Energy-efficient and environmentally friendly production of starch-rich duckweed biomass using nitrogen-limited cultivation. J Clean Prod 251: 119726. https://doi.org/10.1016/j.jclepro.2019.119726
|
| [20] |
Ma Y, Zhu M, Yu C, et al. (2018) Large-scale screening and characterisation of Lemna aequinoctialis and Spirodela polyrhiza strains for starch production. Plant Biol 20: 357-364. https://doi.org/10.1111/plb.12679
|
| [21] |
Sowinski EE, Gilbert S, Lam E, et al. (2019) Linkage structure of cell-wall polysaccharides from three duckweed species. Carbohydr Polym 223: 115119. https://doi.org/10.1016/j.carbpol.2019.115119
|
| [22] |
Pagliuso D, Grandis A, Lam E, et al. (2021) High saccharification, low lignin, and high sustainability potential make duckweeds adequate as bioenergy feedstocks. Bioenergy Res 14: 1082-1092. https://doi.org/10.1007/s12155-020-10211-x
|
| [23] | Ren H, Jiang N, Wang T, et al. (2018) Enhanced biogas production in the duckweed anaerobic digestion process. J Energy Resour Technol 140. https://doi.org/10.1115/1.4039782 |
| [24] |
Kogel KH, Franken P, Hückelhoven R (2006) Endophyte or parasite-what decides?. Curr Opin Plant Biol 9: 358-363. https://doi.org/10.1016/j.pbi.2006.05.001
|
| [25] |
Nutaratat P, Srisuk N, Arunrattiyakorn P, et al. (2014) Plant growth-promoting traits of epiphytic and endophytic yeasts isolated from rice and sugar cane leaves in Thailand. Fungal Biol 118: 683-694. https://doi.org/10.1016/j.funbio.2014.04.010
|
| [26] |
Joubert PM, Doty SL (2018) Endophytic yeasts: Biology, ecology and applications. Endophytes of forest trees . Cham: Springer 3-14. https://doi.org/10.1007/978-3-319-89833-9_1
|
| [27] |
Saimee Y, Duangmal K (2021) Streptomyces spirodelae sp. nov., isolated from duckweed. Int J Syst Evol Microbiol 71: 005106. https://doi.org/10.1099/ijsem.0.005106.
|
| [28] |
Ishizawa H, Kuroda M, Inoue D, et al. (2020) Community dynamics of duckweed-associated bacteria upon inoculation of plant growth-promoting bacteria. FEMS Microbiol Ecol 96: fiaa101. https://doi.org/10.1093/femsec/fiaa101
|
| [29] | Zuki NAAM, Yahya H, Ariffin N, et al. (2022) The classification of duckweed and its bacterial community: A review. Malaysian J Sci 8: 14-26. https://doi.org/10.33102/2022238 |
| [30] | Kittiwongwattana C, Vuttipongchaikij S (2015) Biodiversity of endophytic bacteria isolated from duckweed (Landoltia punctata) and their IAA production. Sci Technol Asia : 1-11. |
| [31] | Bog M, Appenroth KJ, Sree KS (2020) Key to the determination of taxa of Lemnaceae: An update. Nordic J Botany 38. https://doi.org/10.1111/njb.02658 |
| [32] | Ruiz-Barba JL, Maldonado-Barragán A, Jiménez Díaz R (2005) Small-scale total DNA extraction from bacteria and yeast for PCR applications. Anal Biochem 347. https://doi.org/10.1016/j.ab.2005.09.028 |
| [33] |
Kurtzman CP, Robnett CJ (1998) Identification and phylogeny of ascomycetous yeasts from analysis of nuclear large subunit (26S) ribosomal DNA partial sequences. Antonie van Leeuwenhoek 73: 331-371. https://doi.org/10.1023/A:1001761008817
|
| [34] |
Altschul SF, Madden TL, Schäffer AA, et al. (1997) Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res 25: 3389-3402. https://doi.org/10.1093/nar/25.17.3389
|
| [35] |
Vu D, Groenewald M, Szöke S, et al. (2016) DNA barcoding analysis of more than 9 000 yeast isolates contributes to quantitative thresholds for yeast species and genera delimitation. Stud Mycol 85: 91-105. https://doi.org/10.1016/j.simyco.2016.11.007
|
| [36] |
Shannon CE (2001) A mathematical theory of communication. Mob Comput Commun Rev 5: 3-55. https://doi.org/10.1002/j.1538-7305.1948.tb01338.x
|
| [37] |
Spellerberg IF, Fedor PJ (2003) A tribute to Claude Shannon (1916–2001) and a plea for more rigorous use of species richness, species diversity and the ‘Shannon–Wiener’Index. Glob Ecol Biogeogr 12: 177-179. https://doi.org/10.1046/j.1466-822X.2003.00015.x
|
| [38] |
Simpson EH (1949) Measurement of diversity. Nature 163: 688-688. https://doi.org/doi.org/10.1038/163688a0
|
| [39] |
Colwell RK, Chao A, Gotelli NJ, et al. (2012) Models and estimators linking individual-based and sample-based rarefaction, extrapolation and comparison of assemblages. Plant Ecol 5: 3-21. https://doi.org/10.1093/jpe/rtr044
|
| [40] | Hammer Ø, Harper DA, Ryan PD (2001) PAST: Paleontological statistics software package for education and data analysis. Palaeontol Electron 4: 9. |
| [41] |
Nutaratat P, Amsri W, Srisuk N, et al. (2015) Indole-3-acetic acid production by newly isolated red yeast Rhodosporidium paludigenum. J Gen Appl Microbiol 61: 1-9. https://doi.org/10.2323/jgam.61.1
|
| [42] |
Louden BC, Haarmann D, Lynne AM (2011) Use of blue agar CAS assay for siderophore detection. J Microbiol Biol Educ 12: 51-53. https://doi.org/10.1128/jmbe.v12i1.249
|
| [43] |
Fomina MA, Alexander IJ, Colpaert JV, et al. (2005) Solubilization of toxic metal minerals and metal tolerance of mycorrhizal fungi. Soil Biol Biochem 37: 851-866. https://doi.org/10.1016/j.soilbio.2004.10.013
|
| [44] | Bibi F, Ali Z (2013) Measurement of diversity indices of avian communities at Taunsa Barrage Wildlife Sanctuary, Pakistan. J Anim Plant Sci 23: 469-474. |
| [45] |
Glushakova A, Kachalkin A (2017) Endophytic yeasts in Malus domestica and Pyrus communis fruits under anthropogenic impact. Microbiology 86: 128-135. https://doi.org/10.1134/S0026261716060102
|
| [46] |
Isaeva O, Glushakova A, Garbuz S, et al. (2010) Endophytic yeast fungi in plant storage tissues. Biol Bull Rev 37: 26-34. https://doi.org/10.1134/S1062359010010048
|
| [47] |
Limtong S, Koowadjanakul N (2012) Yeasts from phylloplane and their capability to produce indole-3-acetic acid. World J Microbiol 28: 3323-3335. https://doi.org/10.1007/s11274-012-1144-9
|
| [48] |
Khunnamwong P, Jindamorakot S, Limtong S (2018) Endophytic yeast diversity in leaf tissue of rice, corn and sugarcane cultivated in Thailand assessed by a culture-dependent approach. Fungal Biol 122: 785-799. https://doi.org/10.1016/j.funbio.2018.04.006
|
| [49] | Matos ÍTSR, de Souza VA, D'Angelo GdR, et al. (2021) Yeasts with fermentative potential associated with fruits of camu-camu (Myrciaria dubia, Kunth) from North of Brazilian Amazon. Sci World J 2021. https://doi.org/10.1155/2021/9929059 |
| [50] |
Vaz AB, Mota RC, Bomfim MRQ, et al. (2009) Antimicrobial activity of endophytic fungi associated with Orchidaceae in Brazil. Can J Microbiol 55: 1381-1391. https://doi.org/10.1139/W09-101
|
| [51] | Islam F, Salam MA, Rahman MA, et al. (2021) Plant endophytic yeasts Pichia fermentans and Meyerozyma caribbica improve growth, biochemical composition, haematological parameters and morphology of internal organs of premature Barbonymus gonionotus. Aquac Res 19: 100575. https://doi.org/10.1016/j.aqrep.2020.100575 |
| [52] |
de Lima Targino HM, Silva VSL, Escobar IEC, et al. (2022) Maize-associated Meyerozyma from the Brazilian semiarid region are effective plant growth-promoting yeasts. Rhizosphere : 100538. https://doi.org/10.1016/j.rhisph.2022.100538
|
| [53] |
Ling L, Li Z, Jiao Z, et al. (2019) Identification of novel endophytic yeast strains from tangerine peel. Curr Microbiol 76: 1066-1072. https://doi.org/10.1007/s00284-019-01721-9
|
| [54] |
Bura R, Vajzovic A, Doty SL (2012) Novel endophytic yeast Rhodotorula mucilaginosa strain PTD3 I: production of xylitol and ethanol. J Ind Microbiol Biotechnol 39: 1003-1011. https://doi.org/10.1007/s10295-012-1109-x
|
| [55] |
Ramirez-Castrillon M, Usman LM, Silva-Bedoya LM, et al. (2019) Dominant yeasts associated to mango (Mangifera indica) and rose apple (Syzygium malaccense) fruit pulps investigated by culture-based methods. An Acad Bras Cienc 91. https://doi.org/10.1590/0001-3765201920190052
|
| [56] |
Imanishi Y, Jindamorakot S, Mikata K, et al. (2008) Two new ascomycetous anamorphic yeast species related to Candida friedrichii—Candida jaroonii sp. nov., and Candida songkhlaensis sp. nov.—isolated in Thailand. Antonie van Leeuwenhoek 94: 267-276. https://doi.org/10.1007/s10482-008-9242-2
|
| [57] |
Limtong S, Kaewwichian R (2015) The diversity of culturable yeasts in the phylloplane of rice in Thailand. Ann Microbiol 65: 667-675. https://doi.org/10.1007/s13213-014-0905-0
|
| [58] |
Kachalkin AV, Yurkov AM (2012) Yeast communities in Sphagnum phyllosphere along the temperature-moisture ecocline in the boreal forest-swamp ecosystem and description of Candida sphagnicola sp. nov. Antonie van Leeuwenhoek 102: 29-43. https://doi.org/10.1007/s10482-012-9710-6
|
| [59] |
Limtong S, Kaewwichian R, Yongmanitchai W, et al. (2014) Diversity of culturable yeasts in phylloplane of sugarcane in Thailand and their capability to produce indole-3-acetic acid. World J Microbiol 30: 1785-1796. https://doi.org/10.1007/s11274-014-1602-7
|
| [60] |
Hong SG, Chun J, Oh HW, et al. (2001) Metschnikowia koreensis sp. nov., a novel yeast species isolated from flowers in Korea. Int J Syst Evol Microbiol 51: 1927-1931. https://doi.org/10.1099/00207713-51-5-1927
|
| [61] |
Camargo FP, Araujo ACV, de Moraes EM, et al. (2016) A comparison between cactophilic yeast communities isolated from Cereus hildmannianus and Praecereus euchlorus necrotic cladodes. Fungal Biol 120: 1175-1183. https://doi.org/10.1016/j.funbio.2016.06.003
|
| [62] |
Kaewwichian R, Yongmanitchai W, Kawasaki H, et al. (2012) Metschnikowia saccharicola sp. nov. and Metschnikowia lopburiensis sp. nov., two novel yeast species isolated from phylloplane in Thailand. Antonie van Leeuwenhoek 102: 743-751. https://doi.org/10.1007/s10482-012-9774-3
|
| [63] |
Sibirny A Non-conventional yeasts: from basic research to application: Springer (2019). https://doi.org/10.1007/978-3-030-21110-3
|
| [64] |
Lopes MR, Lara CA, Moura ME, et al. (2018) Characterisation of the diversity and physiology of cellobiose-fermenting yeasts isolated from rotting wood in Brazilian ecosystems. Fungal Biol 122: 668-676. https://doi.org/10.1016/j.funbio.2018.03.008
|
| [65] |
Suzuki M, Nakase T, Komagata K (1994) Candida stellimalicola, a new species of anamorphic yeast isolated from star apple in Thailand. J Gen Appl Microbiol 40: 115-121. https://doi.org/10.2323/JGAM.40.115
|
| [66] |
da Cunha T, Ferraz LP, Wehr PP, et al. (2018) Antifungal activity and action mechanisms of yeasts isolates from citrus against Penicillium italicum. Int J Food Microbiol 276: 20-27. https://doi.org/10.1016/j.ijfoodmicro.2018.03.019
|
| [67] |
Kanpiengjai A, Chui-Chai N, Chaikaew S, et al. (2016) Distribution of tannin-'tolerant yeasts isolated from Miang, a traditional fermented tea leaf (Camellia sinensis var. assamica) in northern Thailand. Int J Food Microbiol 238: 121-131. https://doi.org/10.1016/j.ijfoodmicro.2016.08.044
|
| [68] |
Zhang J, Shang Y, Chen J, et al. (2021) Diversity of non-Saccharomyces yeasts of grape berry surfaces from representative Cabernet Sauvignon vineyards in Henan Province, China. FEMS Microbiol Lett 368: fnab142. https://doi.org/10.1093/femsle/fnab142
|
| [69] |
Freitas LF, Batista TM, Santos AR, et al. (2020) Yeast communities associated with cacti in Brazil and the description of Kluyveromyces starmeri sp. nov. based on phylogenomic analyses. Yeast 37: 625-637. https://doi.org/10.1002/yea.3528
|
| [70] |
Nakase T, Jindamorakot S, Am-In S, et al. (2009) Candida nonsorbophila sp. nov., a new ascomycetous yeast species isolated in Thailand. FEMS Yeast Res 9: 663-667. https://doi.org/10.1111/j.1567-1364.2009.00512.x
|
| [71] |
Maciel NO, Santos AR, Felix CR, et al. (2021) Wickerhamiella martinezcruziae fa, sp. nov., a yeast species isolated from tropical habitats. Int J Syst Evol Microbiol 71: 005092. https://doi.org/10.1099/ijsem.0.005092
|
| [72] |
Li AH, Yuan FX, Groenewald M, et al. (2020) Diversity and phylogeny of basidiomycetous yeasts from plant leaves and soil: proposal of two new orders, three new families, eight new genera and one hundred and seven new species. Stud Mycol 96: 17-140. https://doi.org/10.1016/j.simyco.2020.01.002
|
| [73] |
Into P, Pontes A, Sampaio JP, et al. (2020) Yeast diversity associated with the phylloplane of corn plants cultivated in Thailand. Microorganisms 8: 80. https://doi.org/10.3390/microorganisms8010080
|
| [74] |
Osman ME, Abdel-Razik AB, Zaki KI, et al. (2022) Isolation, molecular identification of lipid-producing Rhodotorula diobovata: optimization of lipid accumulation for biodiesel production. J Genet Eng Biotechnol 20: 1-15. https://doi.org/10.1186/s43141-022-00304-9
|
| [75] |
Saluja P, Prasad GS (2007) Debaryomyces singareniensis sp. nov., a novel yeast species isolated from a coal mine soil in India. FEMS Yeast Res 7: 482-488. https://doi.org/10.1111/j.1567-1364.2006.00182.x
|
| [76] |
Limtong S, Srisuk N, Yongmanitchai W, et al. (2005) Pichia thermomethanolica sp. nov., a novel thermotolerant, methylotrophic yeast isolated in Thailand. Int J Syst Evol Microbiol 55: 2225-2229. https://doi.org/10.1099/ijs.0.63712-0
|
| [77] |
Lorliam W, Akaracharanya A, Suzuki M, et al. (2013) Diversity and fermentation products of xylose-utilizing yeasts isolated from buffalo feces in Thailand. Microbes Environ 28: 354-360. https://doi.org/10.1264/jsme2.me13023
|
| [78] |
Sakpuntoon V, Péter G, Groenewald M, et al. (2022) Description of Crinitomyces reliqui gen. nov., sp. nov. and reassignment of Trichosporiella flavificans and Candida ghanaensis to the genus Crinitomyces. J Fungi 8: 224. https://doi.org/10.3390/jof8030224
|
| [79] |
Rodrigues de Miranda L (1978) A new genus: Sporopachydermia. Antonie van Leeuwenhoek 44: 439-450. https://doi.org/10.1007/BF00394320
|
| [80] |
Perpetuini G, Rossetti AP, Battistelli N, et al. (2021) Adhesion properties, biofilm forming potential, and susceptibility to disinfectants of contaminant wine yeasts. Microorganisms 9: 654. https://doi.org/10.3390/microorganisms9030654
|
| [81] |
Camilo S, Chandra M, Branco P, et al. (2022) Wine microbial consortium: Seasonal sources and vectors linking vineyard and winery environments. Ferment 8: 324. https://doi.org/10.3390/fermentation8070324
|
| [82] |
Kurtzman CP (2007) New anamorphic yeast species: Candida infanticola sp. nov., Candida polysorbophila sp. nov., Candida transvaalensis sp. nov. and Trigonopsis californica sp. nov. Antonie Van Leeuwenhoek 92: 221-231. https://doi.org/10.1007/s10482-007-9150-x
|
| [83] | Kolochani MK, Mousavi SM, Zamani I, et al. (2022) Isolation and identification of lactic acid bacteria and yeasts with probiotic ability from the intestine of gilthead seabream. Iran Vet J 18: 74-86. https://doi.org/10.22055/IVJ.2022.353171.2482 |
| [84] |
Monapathi ME, Bezuidenhout CC, James Rhode OH (2020) Aquatic yeasts: diversity, characteristics and potential health implications. J Water Health 18: 91-105. https://doi.org/10.2166/wh.2020.270
|
| [85] |
Suh SO, Blackwell M, Kurtzman CP, et al. (2006) Phylogenetics of Saccharomycetales, the ascomycete yeasts. Mycologia 98: 1006-1017. https://doi.org/10.3852/mycologia.98.6.1006
|
| [86] |
Kim J, Sudbery P (2011) Candida albicans, a major human fungal pathogen. J Microbiol 49: 171-177. https://doi.org/10.1007/s12275-011-1064-7
|
| [87] |
Brilhante RSN, Sales JA, da Silva MLQ, et al. (2018) Antifungal susceptibility and virulence of Candida parapsilosis species complex: an overview of their pathogenic potential. J Med Microbiol 67: 903-914. https://doi.org/10.1099/jmm.0.000756
|
| [88] |
Bertini A, De Bernardis F, Hensgens LA, et al. (2013) Comparison of Candida parapsilosis, Candida orthopsilosis, and Candida metapsilosis adhesive properties and pathogenicity. Int J Med Microbiol 303: 98-103. https://doi.org/10.1016/j.ijmm.2012.12.006
|
| [89] |
Jensen RH, Arendrup MC (2011) Candida palmioleophila: characterization of a previously overlooked pathogen and its unique susceptibility profile in comparison with five related species. J Clin Microbiol 49: 549-556. https://doi.org/10.1128/JCM.02071-10
|
| [90] |
Ann Chai LY, Denning DW, Warn P (2010) Candida tropicalis in human disease. Crit Rev Microbiol 36: 282-298. https://doi.org/10.3109/1040841X.2010.489506
|
| [91] |
Al-Obaid K, Ahmad S, Joseph L, et al. (2018) Lodderomyces elongisporus: a bloodstream pathogen of greater clinical significance. New Microbes New Infect 26: 20-24. https://doi.org/10.1016/j.nmni.2018.07.004
|
| [92] |
Ioannou P, Papakitsou I (2020) Kodamaea ohmeri infections in humans: A systematic review. Mycoses 63: 636-643. https://doi.org/10.1111/myc.13094
|
| [93] |
Glushakova A, Kachalkin A, Zheltikova T, et al. (2015) Yeasts associated with wind-pollinated plants—leading pollen allergens in Central Russia. Microbiology 84: 722-725. https://doi.org/10.1134/S0026261715050082
|
| [94] |
Yalçın HT, Fındık B, Terzi Y, et al. (2021) Isolation and molecular identification of industrially important enzyme producer yeasts from tree barks and fruits. Arch Microbiol 203: 1079-1088. https://doi.org/10.1007/s00203-020-02104-6
|
| [95] |
Prakash A, Randhawa HS, Khan ZU, et al. (2018) Environmental distribution of Cryptococcus species and some other yeast-like fungi in India. Mycoses 61: 305-313. https://doi.org/10.1111/myc.12741
|
| [96] |
Boonmak C, Khunnamwong P, Limtong S (2020) Yeast communities of primary and secondary peat swamp forests in southern Thailand. Antonie Van Leeuwenhoek 113: 55-69. https://doi.org/10.1007/s10482-019-01317-0
|
| [97] |
Vieira NM, Ventorim RZ, de Moura Ferreira MA, et al. (2020) Insights into oleaginous phenotype of the yeast Papiliotrema laurentii. Fungal Genet Biol 144: 103456. https://doi.org/10.1016/j.fgb.2020.103456
|
| [98] |
Cloete KJ, Valentine AJ, Stander MA, et al. (2009) Evidence of symbiosis between the soil yeast Cryptococcus laurentii and a sclerophyllous medicinal shrub, Agathosma betulina (Berg.) Pillans. Microb Ecol 57: 624-632. https://doi.org/10.1007/s00248-008-9457-9
|
| [99] |
Wang G, Liu L, Liang W (2018) Single cell oil production from hydrolysates of inulin by a newly isolated yeast Papiliotrema laurentii AM113 for biodiesel making. Biotechnol Appl Biochem 184: 168-181. https://doi.org/10.1007/s12010-017-2538-9
|
| [100] |
Kachalkin A, Glushakova A, Streletskii R (2022) Diversity of endophytic yeasts from agricultural fruits positive for phytohormone iaa production. BioTech (Basel) 11: 38. https://doi.org/10.3390/biotech11030038
|
| [101] |
Gai CS, Lacava PT, Maccheroni Jr W, et al. (2009) Diversity of endophytic yeasts from sweet orange and their localization by scanning electron microscopy. J Basic Microbiol 49: 441-451. https://doi.org/10.1002/yea.3528
|
| [102] |
Kaewkrajay C, Putchakarn S, Limtong S (2021) Cultivable yeasts associated with marine sponges in the Gulf of Thailand, South China Sea. Antonie van Leeuwenhoek 114: 253-274. https://doi.org/10.1007/s10482-021-01518-6
|
| [103] |
Srisuk N, Nutaratat P, Surussawadee J, et al. (2019) Yeast communities in sugarcane phylloplane. Microbiology 88: 353-369. https://doi.org/10.1134/S0026261719030135
|
| [104] |
Singh A, Chisti Y, Banerjee UC (2011) Production of carbonyl reductase by Metschnikowia koreensis. Bioresour Technol 102: 10679-10685. https://doi.org/10.1016/j.biortech.2011.09.032
|
| [105] |
Jaiboon K, Lertwattanasakul N, Limtong P, et al. (2016) Yeasts from peat in a tropical peat swamp forest in Thailand and their ability to produce ethanol, indole-3-acetic acid and extracellular enzymes. Mycol Prog 15: 755-770. https://doi.org/10.1007/s11557-016-1205-9
|
| [106] | Tan C, Wang L, Xue Y, et al. (2018) Purification and molecular characterization of a Metschnikowia saccharicola killer toxin lethal to a crab pathogenic yeast. FEMS Microbiol Lett 365. https://doi.org/10.1093/femsle/fny038 |
| [107] |
Sousa-Silva M, Vieira D, Soares P, et al. (2021) Expanding the knowledge on the skillful yeast Cyberlindnera jadinii. J Fungi 7: 36. https://doi.org/10.3390/jof7010036
|
| [108] | Nguyen MT, Ranamukhaarachchi SL, Hannaway DB (2011) Efficacy of antagonist strains of Bacillus megaterium, Enterobacter cloacae, Pichia guilliermondii and Candida ethanolica against bacterial wilt disease of tomato. J Phytol 3. |
| [109] |
Agrawal T, Kotasthane AS (2012) Chitinolytic assay of indigenous Trichoderma isolates collected from different geographical locations of Chhattisgarh in Central India. Springerplus 1: 1-10. https://doi.org/10.1186/2193-1801-1-73
|
| [110] |
Vernekar JV, Ghatge MS, Deshpande VV (1999) Alkaline protease inhibitor: a novel class of antifungal proteins against phytopathogenic fungi. Biochem Biophys Res Commun 262: 702-707. https://doi.org/10.1006/bbrc.1999.1269
|
| [111] |
Petkova M, Petrova S, Spasova-Apostolova V, et al. (2022) Tobacco plant growth-promoting and antifungal activities of three endophytic yeast strains. Plants 11: 751. https://doi.org/10.3390/plants11060751
|
| [112] | Nimsi KA, Manjusha K, Kathiresan K, et al. (2022) Plant growth-promoting yeasts (PGPY), the latest entrant for use in sustainable agriculture: a review. J Appl Microbiol 134. https://doi.org/10.1093/jambio/lxac088 |
| [113] |
Gizaw B, Tsegay Z, Tefera G, et al. (2017) Phosphate solubilizing fungi isolated and characterized from teff rhizosphere soil collected from north showa and gojam, Ethiopia. J Fertil Pestic 8. https://doi.org/10.4172/2471-2728.1000180
|
| [114] | Nakayan P, Shen FT, Hung MH, et al. (2009) Effectiveness of Pichia sp. CC1 in decreasing chemical fertilization requirements of garden lettuce in pot experiments. J Food Ag-Ind . |
 microbiol-09-03-026-s001.pdf microbiol-09-03-026-s001.pdf |
 |




Figures(5) / Tables(2)

Napapohn Kajadpai, Jirameth Angchuan, Pannida Khunnamwong, Nantana Srisuk. Diversity of duckweed (Lemnaceae) associated yeasts and their plant growth promoting characteristics[J]. AIMS Microbiology, 2023, 9(3): 486-517. doi: 10.3934/microbiol.2023026







 DownLoad:
DownLoad: