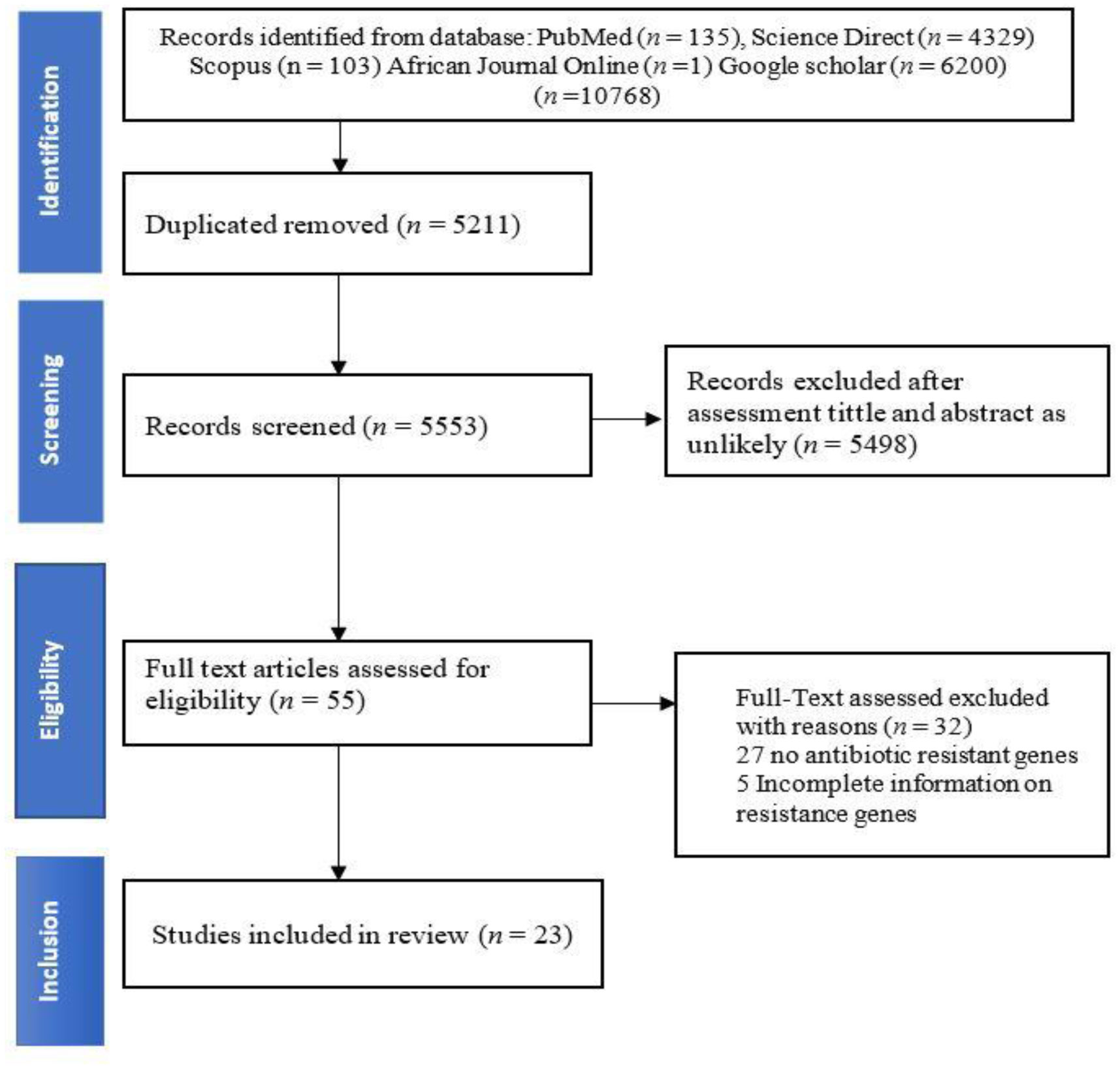
This is a systematic review and meta-analysis that evaluated the prevalence of Escherichia coli antibiotic-resistant genes (ARGs) in animals, humans, and the environment in South Africa. This study followed Preferred Reporting Items for Systematic Reviews and Meta-analyses (PRISMA) guidelines to search and use literature published between 1 January 2000 to 12 December 2021, on the prevalence of South African E. coli isolates' ARGs. Articles were downloaded from African Journals Online, PubMed, ScienceDirect, Scopus, and Google Scholar search engines. A random effects meta-analysis was used to estimate the antibiotic-resistant genes of E. coli in animals, humans, and the environment. Out of 10764 published articles, only 23 studies met the inclusion criteria. The obtained results indicated that the pooled prevalence estimates (PPE) of E. coli ARGs was 36.3%, 34.4%, 32.9%, and 28.8% for blaTEM-M-1, ampC, tetA, and blaTEM, respectively. Eight ARGs (blaCTX-M, blaCTX-M-1, blaTEM, tetA, tetB, sul1, sulII, and aadA) were detected in humans, animals and the environmental samples. Human E. coli isolate samples harboured 38% of the ARGs. Analyzed data from this study highlights the occurrence of ARGs in E. coli isolates from animals, humans, and environmental samples in South Africa. Therefore, there is a necessity to develop a comprehensive “One Health” strategy to assess antibiotics use in order to understand the causes and dynamics of antibiotic resistance development, as such information will enable the formulation of intervention strategies to stop the spread of ARGs in the future.
Citation: Tsepo Ramatla, Mpho Tawana, Kgaugelo E. Lekota, Oriel Thekisoe. Antimicrobial resistance genes of Escherichia coli, a bacterium of “One Health” importance in South Africa: Systematic review and meta-analysis[J]. AIMS Microbiology, 2023, 9(1): 75-89. doi: 10.3934/microbiol.2023005
This is a systematic review and meta-analysis that evaluated the prevalence of Escherichia coli antibiotic-resistant genes (ARGs) in animals, humans, and the environment in South Africa. This study followed Preferred Reporting Items for Systematic Reviews and Meta-analyses (PRISMA) guidelines to search and use literature published between 1 January 2000 to 12 December 2021, on the prevalence of South African E. coli isolates' ARGs. Articles were downloaded from African Journals Online, PubMed, ScienceDirect, Scopus, and Google Scholar search engines. A random effects meta-analysis was used to estimate the antibiotic-resistant genes of E. coli in animals, humans, and the environment. Out of 10764 published articles, only 23 studies met the inclusion criteria. The obtained results indicated that the pooled prevalence estimates (PPE) of E. coli ARGs was 36.3%, 34.4%, 32.9%, and 28.8% for blaTEM-M-1, ampC, tetA, and blaTEM, respectively. Eight ARGs (blaCTX-M, blaCTX-M-1, blaTEM, tetA, tetB, sul1, sulII, and aadA) were detected in humans, animals and the environmental samples. Human E. coli isolate samples harboured 38% of the ARGs. Analyzed data from this study highlights the occurrence of ARGs in E. coli isolates from animals, humans, and environmental samples in South Africa. Therefore, there is a necessity to develop a comprehensive “One Health” strategy to assess antibiotics use in order to understand the causes and dynamics of antibiotic resistance development, as such information will enable the formulation of intervention strategies to stop the spread of ARGs in the future.

| [1] |
Jang J, Hur HG, Sadowsky MJ, et al. (2017) Environmental Escherichia coli: ecology and public health implications—a review. J Appl Microbiol 123: 570-581. https://doi.org/10.1111/jam.13468 
|
| [2] | Meng J, LeJeune JT, Zhao T (2012) Enterohemorrhagic Escherichia coli. Food Microbiol: Fundamentals and frontiers . ASM Press 287-309. https://doi.org/10.1128/9781555818463.ch12 |
| [3] |
Kaper JB, Nataro JP, Mobley HL (2004) Pathogenic Escherichia coli. Nat Rev Microbiol 2: 123-140. https://doi.org/10.1038/nrmicro818
|
| [4] |
Sonola VS, Katakweba A, Misinzo G, et al. (2022) Molecular epidemiology of antibiotic resistance genes and virulence factors in multidrug-resistant Escherichia coli isolated from rodents, humans, chicken, and household soils in Karatu, Northern Tanzania. Int J Environ Res Public Health 19: 5388. https://doi.org/10.3390/ijerph19095388
|
| [5] |
Nobili G, Franconieri I, La Bella G, et al. (2017) Prevalence of verocytotoxigenic Escherichia coli strains isolated from raw beef in southern Italy. Int J Food Microbiol 257: 201-205. https://doi.org/10.1016/j.ijfoodmicro.2017.06.022
|
| [6] | Abd El Shakour EH, Mostafa A (2012) Antimicrobial resistance profiles of Enterobacteriaceae isolated from Rosetta Branch of river Nile, Egypt. World Appl Sci J 19: 1234-1243. https://doi.org/10.5829/idosi.wasj.2012.19.09.2785 |
| [7] |
Peirano G, van Greune CH, Pitout JD (2011) Characteristics of infections caused by extended-spectrum β-lactamase–producing Escherichia coli from community hospitals in South Africa. Diagn Microbiol Infect Dis 69: 449-453. https://doi.org/10.1016/j.diagmicrobio.2010.11.011
|
| [8] |
Racewicz P, Majewski M, Biesiada H (2022) Prevalence and characterisation of antimicrobial resistance genes and class 1 and 2 integrons in multiresistant Escherichia coli isolated from poultry production. Sci Rep 12: 1-13. https://doi.org/10.1038/s41598-022-09996-y
|
| [9] |
Munita JM, Arias CA (2016) Mechanisms of antibiotic resistance. Microbiol Spectr 4: 4-2. https://doi.org/10.1128/microbiolspec.VMBF-0016-2015
|
| [10] |
Esperón F, Sacristán C, Carballo M (2018) Antimicrobial resistance genes in animal manure, manure-amended and nonanthropogenically impacted soils in Spain. Adv Biosci Biotechnol 9: 469-480. https://doi.org/10.4236/abb.2018.99032
|
| [11] |
Muloi DM, Wee BA, McClean DM, et al. (2022) Population genomics of Escherichia coli in livestock-keeping households across a rapidly developing urban landscape. Nat Microbiol 7: 581-589. https://doi.org/10.1038/s41564-022-01079-y
|
| [12] |
Chen X, Zhou L, Tian K, et al. (2013) Metabolic engineering of Escherichia coli: a sustainable industrial platform for bio-based chemical production. Biotechnol Adv 31: 1200-1223. https://doi.org/10.1016/j.biotechadv.2013.02.009
|
| [13] |
Manishimwe R, Moncada PM, Bugarel M, et al. (2021) Antibiotic resistance among Escherichia coli and Salmonella isolated from dairy cattle feces in Texas. Plos One 16: p.e0242390. https://doi.org/10.1371/journal.pone.0242390
|
| [14] |
Su HC, Ying GG, Tao R, et al. (2011) Occurrence of antibiotic resistance and characterization of resistance genes and integrons in Enterobacteriaceae isolated from integrated fish farms in south China. J Environ Monit 13: 3229-3236. https://doi.org/10.1039/c1em10634a
|
| [15] |
Amarasiri M, Sano D, Suzuki S (2020) Understanding human health risks caused by antibiotic resistant bacteria (ARB) and antibiotic resistance genes (ARG) in water environments: Current knowledge and questions to be answered. Crit Rev Environ Sci Technol 50: 2016-2059. https://doi.org/10.1080/10643389.2019.1692611
|
| [16] | Tao S, Chen HLN, Wang T, et al. (2022) The spread of antibiotic resistance genes in vivo model. Can J Infect Dis Med Microbiol 2022. https://doi.org/10.1155/2022/3348695 |
| [17] |
Jian Z, Zeng L, Xu T (2021) Antibiotic resistance genes in bacteria: Occurrence, spread, and control. J Basic Microbiol 61: 1049-1070. https://doi.org/10.1002/jobm.202100201
|
| [18] |
Nys S, Okeke IN, Kariuki S (2004) Antibiotic resistance of faecal Escherichia coli from healthy volunteers from eight developing countries. J Antimicrob Chemother 54: 952-955. https://doi.org/10.1093/jac/dkh448
|
| [19] |
Ekwanzala MD, Dewar JB, Kamika I (2018) Systematic review in South Africa reveals antibiotic resistance genes shared between clinical and environmental settings. Infect Drug Resist 11: 1907. https://doi.org/10.2147/IDR.S170715
|
| [20] |
Ramatla TA, Mphuthi N, Ramaili T (2022) Molecular detection of zoonotic pathogens causing gastroenteritis in humans: Salmonella spp., Shigella spp. and Escherichia coli isolated from Rattus species inhabiting chicken farms in North West Province, South Africa. J S Afr Vet Assoc 93: 1-7. https://doi.org/10.36303/JSAVA.83
|
| [21] |
Adefisoye MA, Okoh AI (2016) Identification and antimicrobial resistance prevalence of pathogenic Escherichia coli strains from treated wastewater effluents in Eastern Cape, South Africa. Microbiologyopen 5: 143-151. https://doi.org/10.1002/mbo3.319
|
| [22] |
Adegoke AA, Madu CE, Aiyegoro OA (2020) Antibiogram and beta-lactamase genes among cefotaxime resistant E. coli from wastewater treatment plant. Antimicrob Resist Infect Control 9: 1-12. https://doi.org/10.1186/s13756-020-0702-4
|
| [23] |
Ateba CN, Bezuidenhout CC (2008) Characterisation of Escherichia coli O157 strains from humans, cattle and pigs in the North-West Province, South Africa. Int J Food Microbiol 128: 181-188. https://doi.org/10.1016/j.ijfoodmicro.2008.08.011
|
| [24] |
Baloyi T, Duvenage S, Du Plessis E, et al. (2022) Multidrug resistant Escherichia coli from fresh produce sold by street vendors in South African informal settlements. Int J Environ Health Res 32: 1513-1528. https://doi.org/10.1080/09603123.2021.1896681
|
| [25] |
Bumunang EW, McAllister TA, Zaheer R (2019) Characterization of non-O157 Escherichia coli from cattle faecal samples in the North-West Province of South Africa. Microorganisms 7: 272. https://doi.org/10.3390/microorganisms7080272
|
| [26] |
Chukwu MO, Abia ALK, Ubomba-Jaswa E (2019) Antibiotic resistance profile and clonality of E. coli isolated from water and paediatric stool samples in the North West province South Africa. J Pure Appl Microbiol 13: 517-530. https://doi.org/10.22207/JPAM.13.1.58
|
| [27] |
Gumede SN, Abia AL, Amoako DG (2021) Analysis of wastewater reveals the spread of diverse Extended-Spectrum β-Lactamase-Producing E. coli strains in uMgungundlovu District, South Africa. Antibiotics 10: 860. https://doi.org/10.3390/antibiotics10070860
|
| [28] |
Igwaran A, Iweriebor BC, Okoh AI (2018) Molecular characterization and antimicrobial resistance pattern of Escherichia coli recovered from wastewater treatment plants in Eastern Cape South Africa. Int J Environ Res Public health 15: 1237. https://doi.org/10.3390/ijerph15061237
|
| [29] | Iweriebor BC, Iwu CJ, Obi LC (2015) Multiple antibiotic resistances among Shiga toxin producing Escherichia coli O157 in feces of dairy cattle farms in Eastern Cape of South Africa. BMC Microbiol 5: 1-9. https://doi.org/10.1186/s12866-015-0553-y |
| [30] |
Iwu CD, du Plessis E, Korsten L (2021) Antibiogram imprints of E. coli O157: H7 recovered from irrigation water and agricultural soil samples collected from two district municipalities in South Africa. Int J Environ Stud 78: 940-953. https://doi.org/10.1080/00207233.2020.1854522
|
| [31] |
Jaja IF, Oguttu J, Jaja CJI (2020) Prevalence and distribution of antimicrobial resistance determinants of Escherichia coli isolates obtained from meat in South Africa. Plos One 15: e0216914. https://doi.org/10.1371/journal.pone.0216914
|
| [32] |
Kubone PZ, Mlisana KP, Govinden U (2020) Antibiotic susceptibility and molecular characterization of uropathogenic Escherichia coli associated with community-acquired urinary tract infections in urban and rural settings in South Africa. Trop Med Infect Dis 5: 176. https://doi.org/10.3390/tropicalmed5040176
|
| [33] |
Mbelle NM, Feldman C, Osei Sekyere J (2019) The resistome, mobilome, virulome and phylogenomics of multidrug-resistant Escherichia coli clinical isolates from Pretoria, South Africa. Sci Rep 9: 1-16. https://doi.org/10.1038/s41598-019-52859-2
|
| [34] |
McIver KS, Amoako DG, Abia ALK (2020) Molecular epidemiology of antibiotic-resistant Escherichia coli from farm-to-fork in intensive poultry production in KwaZulu-Natal, South Africa. Antibiotics 9: 850. https://doi.org/10.3390/antibiotics9120850
|
| [35] |
Mkuhlu NA, Chuks IB, Chikwelu OL (2020) Characterization and antibiotic susceptibility profiles of pathogenic Escherichia coli isolated from diarrhea samples within the Buffalo city metropolitan municipality, eastern Cape, South Africa. Open Microbiol J 14: 321-330. https://doi.org/10.2174/1874434602014010321
|
| [36] |
Montso PK, Mlambo V, Ateba CN (2019) The first isolation and molecular characterization of Shiga Toxin-producing virulent multi-drug resistant atypical enteropathogenic Escherichia coli O177 serogroup from South African Cattle. Front Cell Infect Microbiol 9: 333. https://doi.org/10.3389/fcimb.2019.00333
|
| [37] |
Msolo L, Iweriebor BC, Okoh AI (2020) Antimicrobial resistance profiles of diarrheagenic E. coli (DEC) and Salmonella species recovered from diarrheal patients in selected rural communities of the amathole district municipality, Eastern Cape Province, South Africa. Infect Drug Resist 13: 4615. https://doi.org/10.2147/IDR.S269219
|
| [38] |
Newton-Foot M, Snyman Y, Maloba MRB (2017) Plasmid-mediated mcr-1 colistin resistance in Escherichia coli and Klebsiella spp. clinical isolates from the Western Cape region of South Africa. Antimicrob Resist Infect Control 6: 1-7. https://doi.org/10.1186/s13756-017-0234-8
|
| [39] |
Nontongana N, Sibanda T, Ngwenya E (2014) Prevalence and antibiogram profiling of Escherichia coli pathotypes isolated from the Kat River and the Fort Beaufort abstraction water. Int J Environ Res Public Health 11: 8213-8227. https://doi.org/10.3390/ijerph110808213
|
| [40] |
Nzima B, Adegoke AA, Ofon UA (2020) Resistotyping and extended-spectrum beta-lactamase genes among Escherichia coli from wastewater treatment plants and recipient surface water for reuse in South Africa. New Microbes New Infect 38: 100803. https://doi.org/10.1016/j.nmni.2020.100803
|
| [41] |
Omolajaiye SA, Afolabi KO, Iweriebor BC (2020) Pathotyping and antibiotic resistance profiling of Escherichia coli isolates from children with acute diarrhea in amatole district municipality of Eastern Cape, South Africa. BioMed Res Int 2020. https://doi.org/10.1155/2020/4250165
|
| [42] |
Peirano G, van Greune CH, Pitout JD (2011) Characteristics of infections caused by extended-spectrum β-lactamase–producing Escherichia coli from community hospitals in South Africa. Diagn Microbiol Infect Dis 69: 449-453. https://doi.org/10.1016/j.diagmicrobio.2010.11.011
|
| [43] |
van den Honert MS, Gouws PA, Hoffman LC (2021) Escherichia coli Antibiotic Resistance Patterns from Co-Grazing and Non-Co-Grazing Livestock and Wildlife Species from Two Farms in the Western Cape, South Africa. Antibiotics 10: 618. https://doi.org/10.3390/antibiotics10060618
|
| [44] |
Ramatla T, Tawana M, Onyiche TE (2021) Prevalence of antibiotic resistance in Salmonella serotypes concurrently isolated from the environment, animals, and humans in South Africa: a systematic review and meta-analysis. Antibiotics 10: 1435. https://doi.org/10.3390/antibiotics10121435
|
| [45] |
Onohuean H, Agwu E, Nwodo UU (2022) Systematic review and meta-analysis of environmental Vibrio species–antibiotic resistance. Heliyon 2022: e08845. https://doi.org/10.1016/j.heliyon.2022.e08845
|
| [46] | Mossoro-Kpinde CD, Manirakiza A, Mbecko JR (2015) Antimicrobial resistance of enteric Salmonella in Bangui, central African republic. J Trop Med 2015. https://doi.org/10.1155/2015/483974 |
| [47] |
Bajaj P, Singh NS, Virdi JS (2016) Escherichia coli β-Lactamases: What Really Matters. Front Microbiol 7: 417. https://doi.org/10.3389/fmicb.2016.00417
|
| [48] |
Mokgophi TM, Gcebe N, Fasina F (2021) Antimicrobial resistance profiles of Salmonella isolates on chickens processed and retailed at outlets of the informal market in Gauteng Province, South Africa. Pathogens 10: 273. https://doi.org/10.3390/pathogens10030273
|
| [49] |
Eagar H, Swan G, Van Vuuren M (2012) A survey of antimicrobial usage in animals in South Africa with specific reference to food animals. J S Afr Vet Assoc 83: 1-8.
|
| [50] |
Gao P, Mao D, Luo Y (2012) Occurrence of sulfonamide and tetracycline-resistant bacteria and resistance genes in aquaculture environment. Water Res 46: 2355-2364. https://doi.org/10.4102/jsava.v83i1.16
|
| [51] |
Nguyen F, Starosta AL, Arenz S (2014) Tetracycline antibiotics and resistance mechanisms. Biol Chem 395: 559-575. https://doi.org/10.1515/hsz-2013-0292
|
| [52] |
Rezazadeh M, Baghchesaraei H, Peymani A (2016) Plasmid-mediated quinolone-resistance (qnr) genes in clinical isolates of Escherichia coli collected from several hospitals of Qazvin and Zanjan Provinces, Iran. Osong Public Health Res Perspect 7: 307-312. https://doi.org/10.1016/j.phrp.2016.08.003
|
| [53] |
Taha SA, Omar HH (2019) Characterization of plasmid-mediated qnrA and qnrB genes among Enterobacteriaceae strains: quinolone resistance and ESBL production in Ismailia, Egypt. Egypt J Med Hum Genet 20: 1-7. https://doi.org/10.1186/s43042-019-0026-1
|
| [54] |
Colomer-Lluch M, Jofre J, Muniesa M (2014) Quinolone resistance genes (qnrA and qnrS) in bacteriophage particles from wastewater samples and the effect of inducing agents on packaged antibiotic resistance genes. J Antimicrob Chemother 69: 1265-1274. https://doi.org/10.1093/jac/dkt528
|
| [55] |
Tyson GH, Li C, Hsu CH, et al. (2019) Diverse fluoroquinolone resistance plasmids from retail meat E. coli in the United States. Front Microbiol 10: 2826. https://doi.org/10.3389/fmicb.2019.02826
|
| [56] |
Köser CU, Ellington MJ, Peacock SJ (2014) Whole-genome sequencing to control antimicrobial resistance. Trends Genet 30: 401-407. https://doi.org/10.1016/j.tig.2014.07.003
|
| [57] |
Lekota KE, Bezuidt OKI, Mafofo J (2018) Whole genome sequencing and identification of Bacillus endophyticus and B. anthracis isolated from anthrax outbreaks in South Africa. BMC Microbiol 18: 1-15. https://doi.org/10.1186/s12866-018-1205-9
|
| [58] |
Cooper AL, Low AJ, Koziol AG (2020) Systematic evaluation of whole genome sequence-based predictions of Salmonella serotype and antimicrobial resistance. Front Microbiol 11: 549. https://doi.org/10.3389/fmicb.2020.00549
|
| [59] |
Kumburu HH, Sonda T, van Zwetselaar M (2019) Using WGS to identify antibiotic resistance genes and predict antimicrobial resistance phenotypes in MDR Acinetobacter baumannii in Tanzania. J Antimicrob Chemother 74: 1484-1493. https://doi.org/10.1093/jac/dkz055
|
| [60] | Lerner H, Berg C (2015) The concept of health in One Health and some practical implications for research and education: what is One Health?. Infect Ecol Epidemiol 5: 25300. https://doi.org/10.3402/iee.v5.25300 |
| [61] |
Mackenzie JS, Jeggo M (2019) The One Health approach—Why is it so important?. Tropical Med Infect Dis 4: 88. https://doi.org/10.3390/tropicalmed4020088
|
| [62] |
Ramatla T, Tawana M, Mphuthi MB, et al. (2022) Prevalence and antimicrobial resistance of Campylobacter species in South Africa: A “One Health” approach using systematic review and meta-analysis. Int J Infect Dis 125: 294-304. https://doi.org/10.1016/j.ijid.2022.10.042
|
 microbiol-09-01-005-s001.pdf microbiol-09-01-005-s001.pdf |
 |

Figures(2) / Tables(3)

Tsepo Ramatla, Mpho Tawana, Kgaugelo E. Lekota, Oriel Thekisoe. Antimicrobial resistance genes of Escherichia coli, a bacterium of “One Health” importance in South Africa: Systematic review and meta-analysis[J]. AIMS Microbiology, 2023, 9(1): 75-89. doi: 10.3934/microbiol.2023005







 DownLoad:
DownLoad: