Citation: Le Thanh Dong, Helen V. Espinoza, J. Luis Espinoza. Emerging superbugs: The threat of Carbapenem Resistant Enterobacteriaceae[J]. AIMS Microbiology, 2020, 6(3): 176-182. doi: 10.3934/microbiol.2020012

| [1] |
Thabit AK, Crandon JL, Nicolau DP (2015) Antimicrobial resistance: impact on clinical and economic outcomes and the need for new antimicrobials. Expert Opin Pharmacother 16: 159-177. doi: 10.1517/14656566.2015.993381

|
| [2] |
Kelly AM, Mathema B, Larson EL (2017) Carbapenem-resistant enterobacteriaceae in the community: a scoping review. Int J Antimicrob Agents 50: 127-134. doi: 10.1016/j.ijantimicag.2017.03.012
|
| [3] |
van Duin D, Doi Y (2017) The global epidemiology of carbapenemase-producing enterobacteriaceae. Virulence 8: 460-469. doi: 10.1080/21505594.2016.1222343
|
| [4] | Suay-García B, Pérez-Gracia MT (2019) Present and future of carbapenem-resistant enterobacteriaceae (CRE) infections. Antibiotics (Base 8. |
| [5] |
Friedman ND, Carmeli Y, Walton AL, et al. (2017) Carbapenem-resistant enterobacteriaceae: A strategic roadmap for infection control. Infect Control Hosp Epidemiol 38: 580-594. doi: 10.1017/ice.2017.42
|
| [6] |
Nordmann P, Poirel L (2019) Epidemiology and diagnostics of carbapenem resistance in gram-negative bacteria. Clin Infect Dis 69: S521-S528. doi: 10.1093/cid/ciz824
|
| [7] |
Livorsi DJ, Chorazy ML, Schweizer ML, et al. (2018) A systematic review of the epidemiology of carbapenem-resistant enterobacteriaceae in the United States. Antimicrob Resist Infect Control 7: 55. doi: 10.1186/s13756-018-0346-9
|
| [8] | Dantes R, Mu Y, Belflower R, et al. (2013) National burden of invasive methicillin-resistant Staphylococcus aureus infections, United States, 2011. JAMA Intern Med 173: 1970-1978. |
| [9] |
Young-Xu Y, Kuntz JL, Gerding DN, et al. (2015) Clostridium difficile infection among veterans health administration patients. Infect Control Hosp Epidemiol 36: 1038-1045. doi: 10.1017/ice.2015.138
|
| [10] |
Sievert DM, Ricks P, Edwards JR, et al. (2013) Antimicrobial-resistant pathogens associated with healthcare-associated infections: summary of data reported to the National Healthcare Safety Network at the Centers for Disease Control and Prevention, 2009–2010. Infect Control Hosp Epidemiol 34: 1-14. doi: 10.1086/668770
|
| [11] |
Guh AY, Bulens SN, Mu Y, et al. (2015) Epidemiology of carbapenem-resistant enterobacteriaceae in 7 US communities, 2012–2013. JAMA 314: 1479-1487. doi: 10.1001/jama.2015.12480
|
| [12] |
Bassetti M, Peghin M, Pecori D (2016) The management of multidrug-resistant enterobacteriaceae. Curr Opin Infect Dis 29: 583-594. doi: 10.1097/QCO.0000000000000314
|
| [13] |
Shimasaki T, Seekatz A, Bassis C, et al. (2019) Increased relative abundance of Klebsiella pneumoniae carbapenemase-producing Klebsiella pneumoniae within the gut microbiota is associated with risk of bloodstream infection in long-term acute care hospital patients. Clin Infect Dis 68: 2053-2059. doi: 10.1093/cid/ciy796
|
| [14] |
Christophy R, Osman M, Mallat H, et al. (2017) Prevalence, antibiotic susceptibility and characterization of antibiotic resistant genes among carbapenem-resistant gram-negative bacilli and yeast in intestinal flora of cancer patients in North Lebanon. J Infect Public Health 10: 716-720. doi: 10.1016/j.jiph.2016.10.009
|
| [15] |
Tran DM, Larsson M, Olson L, et al. (2019) High prevalence of colonisation with carbapenem-resistant enterobacteriaceae among patients admitted to Vietnamese hospitals: Risk factors and burden of disease. J Infect 79: 115-122. doi: 10.1016/j.jinf.2019.05.013
|
| [16] |
Espinoza JL (2018) Machine learning for tackling microbiota data and infection complications in immunocompromised patients with cancer. J Intern Med 284: 189-192. doi: 10.1111/joim.12746
|
| [17] |
Cammarota G, Ianiro G, Tilg H, et al. (2017) European consensus conference on faecal microbiota transplantation in clinical practice. Gut 66: 569-580. doi: 10.1136/gutjnl-2016-313017
|
| [18] |
Korach-Rechtman H, Hreish M, Fried C, et al. (2020) Intestinal dysbiosis in carriers of carbapenem-resistant enterobacteriaceae. mSphere 5. doi: 10.1128/mSphere.00173-20
|
| [19] |
Davido B, Batista R, Michelon H, et al. (2017) Is faecal microbiota transplantation an option to eradicate highly drug-resistant enteric bacteria carriage? J Hosp Infect 95: 433-437. doi: 10.1016/j.jhin.2017.02.001
|
| [20] |
Dinh A, Fessi H, Duran C, et al. (2018) Clearance of carbapenem-resistant enterobacteriaceae vs vancomycin-resistant enterococci carriage after faecal microbiota transplant: a prospective comparative study. J Hosp Infect 99: 481-486. doi: 10.1016/j.jhin.2018.02.018
|
| [21] |
Huttner BD, de Lastours V, Wassenberg M, et al. (2019) A 5-day course of oral antibiotics followed by faecal transplantation to eradicate carriage of multidrug-resistant enterobacteriaceae: a randomized clinical trial. Clin Microbiol Infect 25: 830-838. doi: 10.1016/j.cmi.2018.12.009
|
| [22] |
Bilinski J, Grzesiowski P, Sorensen N, et al. (2017) Fecal microbiota transplantation in patients with blood disorders inhibits gut colonization with antibiotic-resistant bacteria: Results of a prospective, single-center study. Clin Infect Dis 65: 364-370. doi: 10.1093/cid/cix252
|
| [23] |
Parpia AS, Ndeffo-Mbah ML, Wenzel NS, et al. (2016) Effects of response to 2014–2015 Ebola outbreak on deaths from Malaria, HIV/AIDS, and Tuberculosis, West Africa. Emerg Infect Dis 22: 433-441. doi: 10.3201/eid2203.150977
|
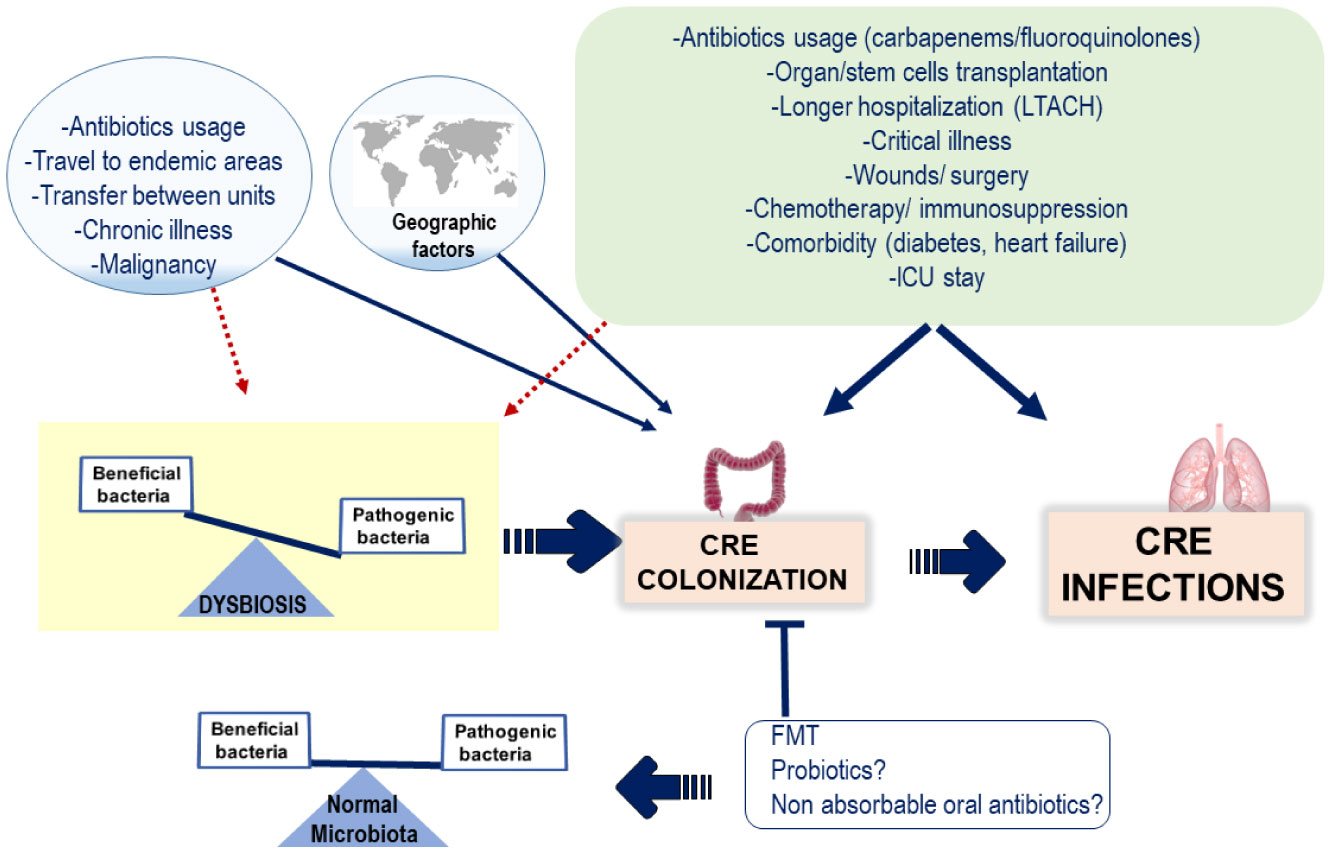
Figures(1)

Le Thanh Dong, Helen V. Espinoza, J. Luis Espinoza. Emerging superbugs: The threat of Carbapenem Resistant Enterobacteriaceae[J]. AIMS Microbiology, 2020, 6(3): 176-182. doi: 10.3934/microbiol.2020012







 DownLoad:
DownLoad: