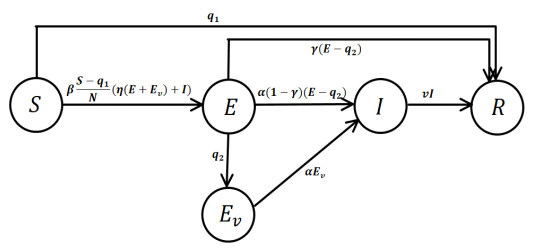
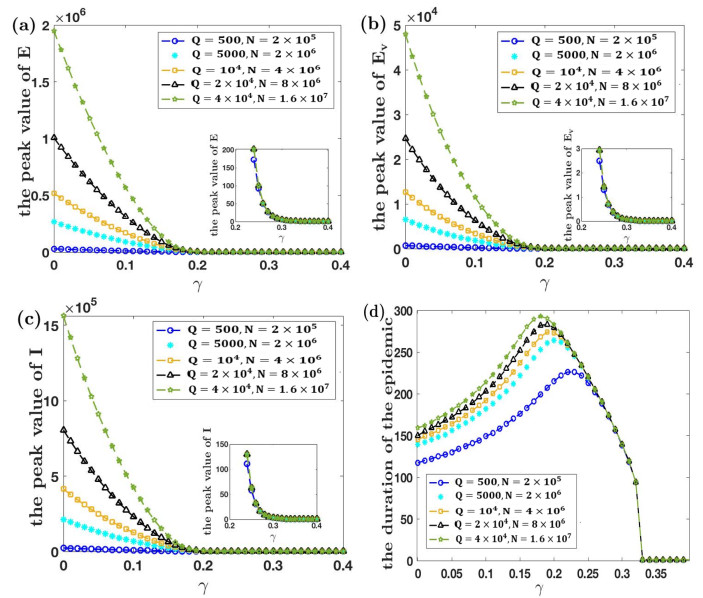
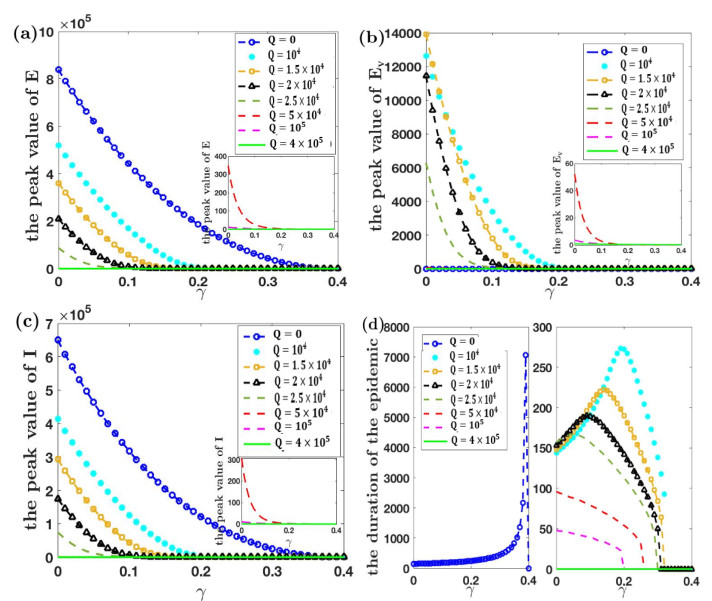
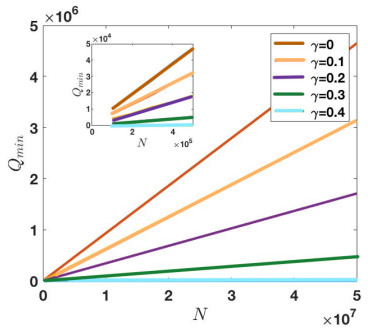
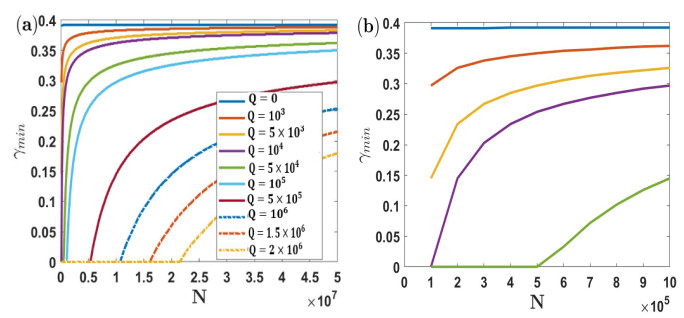
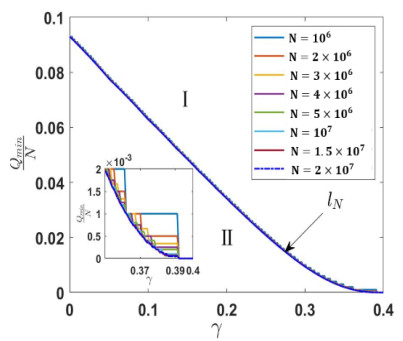
Under the background that asymptomatic virus carriers have infectivity for an infectious disease, we establish a difference equations model with vaccination and virus testing in this paper. Assuming that the vaccine is 100% effective for susceptible people but cannot stop the infectivity of asymptomatic virus carriers, we study how to combine vaccination and virus testing at the beginning of an epidemic to effectively block the spread of infectious disease in different population sizes. By considering the daily processing capacity of the vaccine and daily proportion of testing, the corresponding numerical simulation results are obtained. It is shown that when vaccine availability and virus testing capacity are insufficient, a reasonable combination of the above two measures can slow down or even block the spread of infectious disease. Single virus testing or vaccination can also block the spread of infectious disease, but this requires a lot of manpower, material and financial resources. When the daily proportion of virus testing is fixed, the ratio of the minimum daily processing capacity of vaccines used to block the spread of infectious disease to the corresponding population size is rather stable. It demonstrates that effective protective measures of the same infectious disease in countries and regions with different population sizes can be used as a reference. These results also provide a certain reference for decision makers on how to coordinate vaccines and virus testing resources to curb the spread of such an infectious disease in a certain population size.
Citation: Lili Han, Mingfeng He, Xiao He, Qiuhui Pan. Synergistic effects of vaccination and virus testing on the transmission of an infectious disease[J]. Mathematical Biosciences and Engineering, 2023, 20(9): 16114-16130. doi: 10.3934/mbe.2023719
Under the background that asymptomatic virus carriers have infectivity for an infectious disease, we establish a difference equations model with vaccination and virus testing in this paper. Assuming that the vaccine is 100% effective for susceptible people but cannot stop the infectivity of asymptomatic virus carriers, we study how to combine vaccination and virus testing at the beginning of an epidemic to effectively block the spread of infectious disease in different population sizes. By considering the daily processing capacity of the vaccine and daily proportion of testing, the corresponding numerical simulation results are obtained. It is shown that when vaccine availability and virus testing capacity are insufficient, a reasonable combination of the above two measures can slow down or even block the spread of infectious disease. Single virus testing or vaccination can also block the spread of infectious disease, but this requires a lot of manpower, material and financial resources. When the daily proportion of virus testing is fixed, the ratio of the minimum daily processing capacity of vaccines used to block the spread of infectious disease to the corresponding population size is rather stable. It demonstrates that effective protective measures of the same infectious disease in countries and regions with different population sizes can be used as a reference. These results also provide a certain reference for decision makers on how to coordinate vaccines and virus testing resources to curb the spread of such an infectious disease in a certain population size.

| [1] |
L. Kim, L. McGee, S. Tomczyk, B. Beall, Biological and epidemiological features of antibiotic-resistant streptococcus pneumoniae in pre- and post-conjugate vaccine eras: a united state perspective, Clin. Microbiol. Rev., 29 (2016), 525–552. https://doi.org/10.1128/CMR.00058-15 doi: 10.1128/CMR.00058-15

|
| [2] |
S. K. Saha, B. Hossain, M. Islam, Md Hasanuzzaman, S. Saha, M. Hasan, et al., Epidemiology of invasive pneumococcal disease in Bangladeshi children before introduction of pneumococcal conjugate vaccine, Pediatr. Infect. Dis. J., 35 (2016), 655–661. https://doi.org/10.1097/INF.0000000000001037 doi: 10.1097/INF.0000000000001037
|
| [3] |
H. W. Hethcote, Three basic epidemiological models, Appl. Math. Ecol., 18 (1989), 119–144. https://doi.org/10.1007/978-3-642-61317-3_5 doi: 10.1007/978-3-642-61317-3_5
|
| [4] |
H. W. Hethcote, The mathematics of infectious diseases, SIAM Rev., 42 (2000), 599–653. https://doi.org/10.1137/S0036144500371907 doi: 10.1137/S0036144500371907
|
| [5] |
H. W. Hethcote, P. Van den Driessche, Some epidemiological models with nonlinear Incidence, Journal of Mathematical Biology, 29 (1991), 271–287. https://doi.org/10.1007/BF00160539 doi: 10.1007/BF00160539
|
| [6] |
J. T. F. Lau, X. L. Yang, E. L. Pang, H. Y. Tsui, E. Wong, Y. K. Wing, SARS-Related perceptions in Hong Kong, Emerg. Infect. Dis., 11 (2005), 417–424. https://doi.org/10.3201/eid1103.040675 doi: 10.3201/eid1103.040675
|
| [7] |
L. A. Grohskopf, L. Z. Sokolow, K. R. Broder, E. B. Walter, A. M. Fry, D. B. Jernigan, Prevention and control of seasonal influenza with vaccines: recommendations of the advisory committee on immunization Practices-United States, 2018-19 Influenza Season, MMWR Recomm. Rep., 68 (2019), 1–20. http://dx.doi.org/10.15585/mmwr.rr6803a1 doi: 10.15585/mmwr.rr6803a1
|
| [8] |
D. L. Chao, M. E. Halloran, I. M. Longini, Vaccination strategies for epidemic cholera in Haiti with implications for the developing world, Proc. Natl. Acad. Sci. U. S. A., 108 (2011), 7081–7085. https://doi.org/10.1073/pnas.1102149108 doi: 10.1073/pnas.1102149108
|
| [9] |
J. Q. Li, Z. E. Ma, Y. C. Zhou, Global analysis of SIS epidemic model with a simple vaccination and multiple endemic equilibria, Acta Math. Sci., 26 (2006), 83–93. https://doi.org/10.1016/S0252-9602(06)60029-5 doi: 10.1016/S0252-9602(06)60029-5
|
| [10] |
I. A. Moneim, An SEIR model with infectious latent and a periodic vaccination strategy, Math. Model. Anal., 26 (2021), 236–252. https://doi.org/10.3846/mma.2021.12945 doi: 10.3846/mma.2021.12945
|
| [11] |
C. L. Li, C. H. Li, Dynamics of an epidemic model with imperfect vaccinations on complex networks, J. Phys. A Math. Theor., 53 (2020), 464001. https://doi.org/10.1088/1751-8121/abb9ee doi: 10.1088/1751-8121/abb9ee
|
| [12] |
B. Tang, X. Zhang, Q. Li, N. L. Bragazzi, D. Golemi-Kotra, J. Wu, The minimal COVID-19 vaccination coverage and efficacy to compensate for potential increase of transmission contacts, and increased transmission probability of the emerging strains, BMC Public Health, 22 (2022), 1258. https://doi.org/10.1186/s12889-022-13429-w doi: 10.1186/s12889-022-13429-w
|
| [13] |
Q. Q. Cui, Q. Zhang, Z. P. Qiu, X. Yang, Transmission dynamics of an epidemic model with vaccination, treatment and isolation, Bull. Malaysian Math. Sci. Soc., 42 (2019), 885–896. https://doi.org/10.1007/s40840-017-0519-3 doi: 10.1007/s40840-017-0519-3
|
| [14] |
D. P. Gao, N. J. Huang, S. M. Kang, C. Zhang, Global stability analysis of an SVEIR epidemic model with general incidence rate, Bound. Value Probl., 2018 (2018), 42. https://doi.org/10.1186/s13661-018-0961-7 doi: 10.1186/s13661-018-0961-7
|
| [15] |
G. P. Sahu, J. Dhar, Analysis of an SVEIS epidemic model with partial temporary immunity and saturation incidence rate, Appl. Math. Model., 36 (2012), 908–923. https://doi.org/10.1016/j.apm.2011.07.044 doi: 10.1016/j.apm.2011.07.044
|
| [16] |
J. Liu, K. Wang, Hopf bifurcation of a delayed SIQR epidemic model with constant input and nonlinear incidence rate, Adv. Differ. Equations, 2016 (2016), 168. https://doi.org/10.1186/s13662-016-0899-y doi: 10.1186/s13662-016-0899-y
|
| [17] |
X. Y. Chen, J. D. Cao, J. H. Park, J. Qiu, Stability analysis and estimation of domain of attraction for the endemic equilibrium of an SEIQ epidemic model, Nonlinear Dyn., 87 (2017), 975–985. https://doi.org/10.1007/s11071-016-3092-7 doi: 10.1007/s11071-016-3092-7
|
| [18] |
G. J. Lan, Z. W. Chen, C. J. Wei, S. Zhang, Stationary distribution of a stochastic SIQR epidemic model with saturated incidence and degenerate diffusion, Phys. A Stat. Mech. Appl., 511 (2018), 61–77. https://doi.org/10.1016/j.physa.2018.07.041 doi: 10.1016/j.physa.2018.07.041
|
| [19] |
B. Tang, F. Scarabel, N. L. Bragazzi, Z. McCarthy, M. Glazer, Y. Xiao, et al., De-Escalation by reversing the escalation with a stronger synergistic package of contact tracing, quarantine, isolation and personal protection: Feasibility of preventing a COVID-19 rebound in Ontario, Canada, as a case study, Biol. Basel, 9 (2022), 100. https://doi.org/10.3390/biology9050100 doi: 10.3390/biology9050100
|
| [20] |
L. X. Feng, S. L. Jing, S. K. Hu, D. Wang, H. Huo, Modelling the effects of media coverage and quarantine on the COVID-19 infections in the UK, Math. Biosci. Eng., 17 (2020), 3618–3636. https://doi.org/10.3934/mbe.2020204 doi: 10.3934/mbe.2020204
|
| [21] |
D. A. M. Villela, Imperfect testing of individuals for infectious diseases: Mathematical model and analysis, Commun. Nonliner Sci. Numerical Simul., 46 (2017), 153–160. https://doi.org/10.1016/j.cnsns.2016.10.010 doi: 10.1016/j.cnsns.2016.10.010
|
| [22] |
S. Moore, E. M. Hill, M.J. Tildesley, L. Dyson, M. J. Keeling, Vaccination and non-pharmaceutical interventions for COVID-19: A mathematical modelling study, Lancet Infect. Dis., 21 (2021), 793–802. https://doi.org/10.1016/S1473-3099(21)00143-2 doi: 10.1016/S1473-3099(21)00143-2
|
| [23] |
N. Steyn, M. J. Plank, R. N. Binny, S. C. Hendy, A. Lustig, K. Ridings, A COVID-19 vaccination model for Aotearoa New Zealand, Sci. Rep., 12 (2022), 2720. https://doi.org/10.1038/s41598-022-06707-5 doi: 10.1038/s41598-022-06707-5
|
| [24] |
B. H. Foy, B. Wahl, K. Mehta, A. Shet, G. I. Menon, C. Britto, Comparing COVID-19 vaccine allocation strategies in India: A mathematical modelling study, Int. J. Infect. Dis., 103 (2021), 431–438. https://doi.org/10.1016/j.ijid.2020.12.075 doi: 10.1016/j.ijid.2020.12.075
|
| [25] |
S. A. Rella, Y. A. Kulikova, E. T. Dermitzakis, A. Shet, G. I. Menon, C. Britto, Rates of SARS-CoV-2 transmission and vaccination impact the fate of vaccine-resistant strains, Sci. Rep., 11 (2021), 15729. https://doi.org/10.1038/s41598-021-95025-3 doi: 10.1038/s41598-021-95025-3
|
| [26] |
J. L. Wang, D. Wu, H. Q. Sun, Analysis of an SVIC model with age-dependent infection and asymptomatic carriers, Appl. Anal., 97 (2018), 1467–1495. https://doi.org/10.1080/00036811.2017.1313409 doi: 10.1080/00036811.2017.1313409
|
| [27] |
S. Kurmi, U. Chouhan, A multicompartment mathematical model to study the dynamic behaviour of COVID-19 using vaccination as control parameter, Nonlinear Dyn., 109 (2022), 2185–2201. https://doi.org/10.1007/s11071-022-07591-4 doi: 10.1007/s11071-022-07591-4
|
| [28] |
X. Wang, S. Y. Tang, Y. Chen, X. Feng, Y. Xiao, Z. Xu, When will be the resumption of work in Wuhan and its surrounding areas during COVID-19 epidemic? A data-driven network modeling analysis, Sci. Sinica Math., 50 (2020), 969–978. https://doi.org/10.1360/SSM-2020-0037 doi: 10.1360/SSM-2020-0037
|
| [29] |
T. M. Chen, J. Rui, Q. P. Wang, Z. Zhao, J. Cui, L. Yin, A mathematical model for simulating the phase-based transmissibility of a novel coronavirus, Infect. Dis. Poverty, 9 (2020), 24. https://doi.org/10.1186/s40249-020-00640-3 doi: 10.1186/s40249-020-00640-3
|
| [30] |
Z. F. Yang, Z. Q. Zeng, K. Wang, S. Wong, W. Liang, M. Zanin, Modified SEIR and AI prediction of the epidemics trend of COVID-19 in China under public health interventions, J. Thorac. Dis., 12 (2020), 165–174. https://doi.org/10.21037/jtd.2020.02.64 doi: 10.21037/jtd.2020.02.64
|
| [31] |
R. Pung, C. J. Chiew, B. E. Young, S. Chin, M. Chen, H. E. Clapham, Investigation of three clusters of COVID-19 in Singapore: implications for surveillance and response measures, Lancet, 395 (2020), 1039–1046. https://doi.org/10.1016/S0140-6736(20)30528-6 doi: 10.1016/S0140-6736(20)30528-6
|
| [32] |
W. Guan, Z. Ni, Y. Hu, W. Liang, C. Ou, J. He, Clinical characteristics of coronavirus disease 2019 in China, N. Engl. J. Med., 382 (2020), 1708–1720. https://doi.org/10.1056/NEJMoa2002032 doi: 10.1056/NEJMoa2002032
|
| [33] |
L. L. Han, Q. H. Pan, B. L. Kang, M. F. He, Effects of masks on the transmission of infectious diseases, Adv. Differ. Equations, 2021 (2021), 169. https://doi.org/10.1186/s13662-021-03321-z doi: 10.1186/s13662-021-03321-z
|
| [34] |
P. van den Driessche, Reproduction numbers of infectious disease models, Infect. Dis. Model., 2 (2017), 288–303. https://doi.org/10.1016/j.idm.2017.06.002 doi: 10.1016/j.idm.2017.06.002
|
| [35] | P. van den Driessche, J. Watmough, Further notes on the basic reproduction number, in Mathematical Epidemiology, Springer-Verlag Berlin Heidelberg, 1945 (2008), 159–178. https://doi.org/10.1007/978-3-540-78911-6_6 |
| [36] | National Health Commission of the People's Republic of China: Home: daily Briefing (February 1 to March 18)(2020), 2020. Available from: http://en.nhc.gov.cn/DailyBriefing.html. |
Figures(6) / Tables(1)

Lili Han, Mingfeng He, Xiao He, Qiuhui Pan. Synergistic effects of vaccination and virus testing on the transmission of an infectious disease[J]. Mathematical Biosciences and Engineering, 2023, 20(9): 16114-16130. doi: 10.3934/mbe.2023719







 DownLoad:
DownLoad: