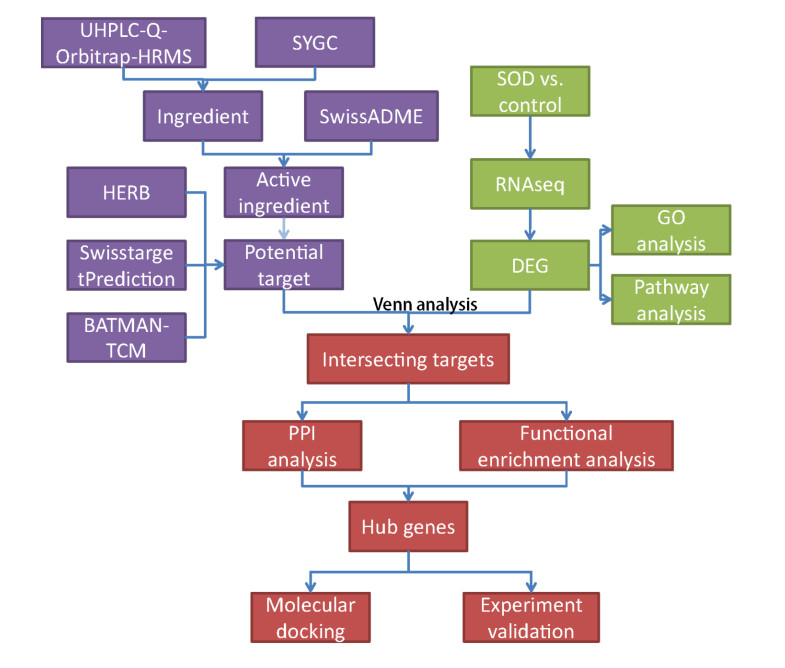
This study explored the chemical and pharmacological mechanisms of Shao Yao Gan Cao decoction (SYGC) in the treatment of Sphincter of Oddi Dysfunction (SOD) through ultra-high-performance liquid chromatography coupled with Quadrupole Exactive-Orbitrap high-resolution mass spectrometry (UHPLC-Q Exactive-Orbitrap HR-MS), network pharmacology, transcriptomics, molecular docking and in vivo experiments. First, we identified that SYGC improves SOD in guinea pigs by increased c-kit expression and decreased inflammation infiltration and ring muscle disorders. Then, a total of 649 SOD differential genes were found through RNA sequencing and mainly enriched in complement and coagulation cascades, the B cell receptor signaling pathway and the NF-kappa B signaling pathway. By combining UHPLC-Q-Orbitrap-HRMS with a network pharmacology study, 111 chemicals and a total of 52 common targets were obtained from SYGC in the treatment of SOD, which is also involved in muscle contraction, the B cell receptor signaling pathway and the complement system. Next, 20 intersecting genes were obtained among the PPI network, MCODE and ClusterOne analysis. Then, the molecular docking results indicated that four active compounds (glycycoumarin, licoflavonol, echinatin and homobutein) and three targets (AURKB, KIF11 and PLG) exerted good binding interactions, which are also related to the B cell receptor signaling pathway and the complement system. Finally, animal experiments were conducted to confirm the SYGC therapy effects on SOD and verify the 22 hub genes using RT-qPCR. This study demonstrates that SYGC confers therapeutic effects against an experimental model of SOD via regulating immune response and inflammation, which provides a basis for future research and clinical applications.
Citation: Yong-hong Hu, Xue-ying Wang, Xi-wen Zhang, Jian Chen, Fu Li. Investigation of the mechanisms and experimental verification of Shao yao gan cao decoction against Sphincter of Oddi Dysfunction via systems pharmacology[J]. Mathematical Biosciences and Engineering, 2022, 19(12): 13374-13398. doi: 10.3934/mbe.2022626
This study explored the chemical and pharmacological mechanisms of Shao Yao Gan Cao decoction (SYGC) in the treatment of Sphincter of Oddi Dysfunction (SOD) through ultra-high-performance liquid chromatography coupled with Quadrupole Exactive-Orbitrap high-resolution mass spectrometry (UHPLC-Q Exactive-Orbitrap HR-MS), network pharmacology, transcriptomics, molecular docking and in vivo experiments. First, we identified that SYGC improves SOD in guinea pigs by increased c-kit expression and decreased inflammation infiltration and ring muscle disorders. Then, a total of 649 SOD differential genes were found through RNA sequencing and mainly enriched in complement and coagulation cascades, the B cell receptor signaling pathway and the NF-kappa B signaling pathway. By combining UHPLC-Q-Orbitrap-HRMS with a network pharmacology study, 111 chemicals and a total of 52 common targets were obtained from SYGC in the treatment of SOD, which is also involved in muscle contraction, the B cell receptor signaling pathway and the complement system. Next, 20 intersecting genes were obtained among the PPI network, MCODE and ClusterOne analysis. Then, the molecular docking results indicated that four active compounds (glycycoumarin, licoflavonol, echinatin and homobutein) and three targets (AURKB, KIF11 and PLG) exerted good binding interactions, which are also related to the B cell receptor signaling pathway and the complement system. Finally, animal experiments were conducted to confirm the SYGC therapy effects on SOD and verify the 22 hub genes using RT-qPCR. This study demonstrates that SYGC confers therapeutic effects against an experimental model of SOD via regulating immune response and inflammation, which provides a basis for future research and clinical applications.

| [1] |
K. R. K. K. Baig, C. M. Wilcox, Translational and clinical perspectives on sphincter of Oddi dysfunction, Clin. Exp. Gastroenterol., 9 (2016), 191–195. https://doi.org/10.2147/CEG.S84018 doi: 10.2147/CEG.S84018

|
| [2] |
P. B. Cotton, G. H. Elta, C. R. Carter, P. J. Pasricha, E. S. Corazziari, I. V. Rome, Gallbladder and Sphincter of Oddi disorders, Gastroenterology, 150 (2016), 1420–1429. https://doi.org/10.1053/j.gastro.2016.02.033 doi: 10.1053/j.gastro.2016.02.033
|
| [3] |
E. Afghani, S. K. Lo, P. S. Covington, B. D. Cash, S. J. Pandol, Sphincter of Oddi function and risk factors for dysfunction, Front. Nutr., 4 (2017), 1. https://doi.org/10.3389/fnut.2017.00001 doi: 10.3389/fnut.2017.00001
|
| [4] |
F. Hinoshita, Y. Ogura, Y. Suzuki, S. Hara, A. Yamada, N. Tanaka, et al., Effect of orally administered shao-yao-gan-cao-tang (Shakuyaku-kanzo-to) on muscle cramps in maintenance hemodialysis patients: A preliminary study, Am. J. Chin. Med., 31 (2003), 445–453. https://doi.org/10.1142/S0192415X03001144 doi: 10.1142/S0192415X03001144
|
| [5] |
T. H. Wu, L. C. Chen, L. L. Yang, Hypouricemic effect and regulatory effects on autonomic function of Shao-Yao Gan-Cao Tang, a Chinese herbal prescription, in asymptomatic hyperuricemic vegetarians, Rheumatol. Int., 28 (2007), 27–31. https://doi.org/10.1007/s00296-007-0385-7 doi: 10.1007/s00296-007-0385-7
|
| [6] |
H. Fujinami, S. Kajiura, J. Nishikawa, T. Ando, T. Sugiyama, The influence of duodenally-delivered Shakuyakukanzoto (Shao Yao Gan Cao Tang) on duodenal peristalsis during endoscopic retrograde cholangiopancreatography: A randomised controlled trial, Chin. Med., 12 (2017), 3. https://doi.org/10.1186/s13020-016-0125-6 doi: 10.1186/s13020-016-0125-6
|
| [7] | M. Chen, F. Li, X. J. Li, Y. P. Hu, B. Gong, X. W. Zhang, et al., Clinical study on the treatment of sphincter of Oddi dysfunction type Ⅱ with Shaoyao Gancao decoction, J. Guangzhou Univ. Tradit. Chin. Med., 38 (2021), 681–686. |
| [8] |
J. Chen, Z. Q. Liang, C. Hu, Y. Gao, Y. K. Wang, J. W. Yang, et al., Protection against peripheral artery disease injury by Ruan Jian Qing Mai formula via metabolic programming, Biotechnol. Appl. Biochem., 68 (2021), 366–380. https://doi.org/10.1002/bab.1934 doi: 10.1002/bab.1934
|
| [9] |
J. Chen, Y. K. Wang, Y. Gao, L. S. Hu, J. W. Yang, J. R. Wang, et al., Protection against COVID-19 injury by qingfei paidu decoction via anti-viral, anti-inflammatory activity and metabolic programming, Biomed. Pharmacother., 129 (2020), 110281. https://doi.org/10.1016/j.biopha.2020.110281 doi: 10.1016/j.biopha.2020.110281
|
| [10] |
M. Yang, J. Sun, Z. Lu, G. Chen, S. Guan, X. Liu, et al., Phytochemical analysis of traditional Chinese medicine using liquid chromatography coupled with mass spectrometry, J. Chromatogr. A, 1216 (2009), 2045–2062. https://doi.org/10.1016/j.chroma.2008.08.097 doi: 10.1016/j.chroma.2008.08.097
|
| [11] |
A. Daina, O. Michielin, V. Zoete, SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules, Sci. Rep., 7 (2017), 42717. https://doi.org/10.1038/srep42717 doi: 10.1038/srep42717
|
| [12] |
S. Fang, L. Dong, L. Liu, J. Guo, L. Zhao, J. Zhang, et al., HERB: A high-throughput experiment- and reference-guided database of traditional Chinese medicine, Nucleic Acids Res., 49 (2021), D1197–D1206. https://doi.org/10.1093/nar/gkaa1063 doi: 10.1093/nar/gkaa1063
|
| [13] |
A. Daina, O. Michielin, V. Zoete, SwissTargetPrediction: updated data and new features for efficient prediction of protein targets of small molecules, Nucleic Acids Res., 47 (2019), W357–W364. https://doi.org/10.1093/nar/gkz382 doi: 10.1093/nar/gkz382
|
| [14] |
Z. Liu, F. Guo, Y. Wang, C. Li, X. Zhang, H. Li, et al., BATMAN-TCM: A bioinformatics analysis tool for molecular mechanism of traditional Chinese medicine, Sci. Rep., 6 (2016), 21146. https://doi.org/10.1038/srep21146 doi: 10.1038/srep21146
|
| [15] |
Y. Zhou, B. Zhou, L. Pache, M. Chang, A. H. Khodabakhshi, O. Tanaseichuk, et al., Metascape provides a biologist-oriented resource for the analysis of systems-level datasets, Nat. Commun., 10 (2019), 1523. https://doi.org/10.1038/s41467-019-09234-6 doi: 10.1038/s41467-019-09234-6
|
| [16] |
G. Su, J. H. Morris, B. Demchak, G. D. Bader, Biological network exploration with Cytoscape 3, Curr. Protoc. Bioinf., 47 (2014), 1–24. https://doi.org/10.1002/0471250953.bi0813s47 doi: 10.1002/0471250953.bi0813s47
|
| [17] |
T. Nepusz, H. Yu, A. Paccanaro, Detecting overlapping protein complexes in protein-protein interaction networks, Nat. Methods, 9 (2012), 471–472. https://doi.org/10.1038/nmeth.1938 doi: 10.1038/nmeth.1938
|
| [18] |
W. D. Leung, S. Sherman, Endoscopic approach to the patient with motility disorders of the bile duct and sphincter of Oddi, Gastrointest. Endosc. Clin., 23 (2013), 405–434. https://doi.org/10.1016/j.giec.2012.12.006 doi: 10.1016/j.giec.2012.12.006
|
| [19] |
G. Y. Zhu, D. D. Jia, Y. Yang, Y. Miao, C. Wang, C. M. Wang, The effect of Shaoyao Gancao decoction on sphincter of Oddi dysfunction in hypercholesterolemic rabbits via protecting the enteric nervous system-interstitial cells of cajal-smooth muscle cells network, J. Inflamm. Res., 14 (2021), 4615–4628. https://doi.org/10.2147/JIR.S326416 doi: 10.2147/JIR.S326416
|
| [20] |
J. Park, D. Jeong, M. Song, B. Kim, Recent advances in anti-metastatic approaches of herbal medicines in 5 major cancers: from traditional medicine to modern drug discovery, Antioxidants, 10 (2021), 527. https://doi.org/10.3390/antiox10040527 doi: 10.3390/antiox10040527
|
| [21] |
Y. Y. Shao, Y. T. Guo, J. P. Gao, J. J. Liu, Z. P. Chang, X. J. Feng, et al., Shaoyao-Gancao decoction relieves visceral hyperalgesia in TNBS-induced postinflammatory irritable bowel syndrome via inactivating transient receptor potential vanilloid type 1 and reducing serotonin synthesis, Evid. Based Complement. Alternat. Med., 2020 (2020), 7830280. https://doi.org/10.1155/2020/7830280 doi: 10.1155/2020/7830280
|
| [22] |
F. Wang, Y. Yang, X. Ji, X. Tao, Y. Wang, C. Wang, Effects of Paeoniflorin on the activity of muscle strips, intracellular calcium ion concentration and Ltype voltagesensitive calcium ion channels in the sphincter of Oddi of hypercholesterolemic rabbits, Mol. Med. Rep., 19 (2019), 5185–5194. https://doi.org/10.3892/mmr.2019.10183 doi: 10.3892/mmr.2019.10183
|
| [23] |
B. Liu, J. Yang, Q. Wen, Y. Li, Isoliquiritigenin, a flavonoid from licorice, relaxes guinea-pig tracheal smooth muscle in vitro and in vivo: role of cGMP/PKG pathway, Eur. J. Pharmacol., 587 (2008), 257–266. https://doi.org/10.1016/j.ejphar.2008.03.015 doi: 10.1016/j.ejphar.2008.03.015
|
| [24] | M. A. Ballal, P. A. Sanford, Physiology of the sphincter of Oddi—the present and the future?—Part 1, Saudi J. Gastroenterol., 6 (2000), 129–146. https://www.saudijgastro.com/text.asp?2000/6/3/129/33475 |
| [25] |
Y. Wang, H. Chang, Y. Zhang, K. Wang, H. Zhang, X. Yan, et al., Endoscopic endoclip papilloplasty preserves sphincter of oddi function, Eur. J. Clin. Invest., 51 (2021), e13408. https://doi.org/10.1111/eci.13408 doi: 10.1111/eci.13408
|
| [26] |
H. Sato, T. Kodama, J. Takaaki, Y. Tatsumi, T. Maeda, S. Fujita, et al., Endoscopic papillary balloon dilatation may preserve sphincter of Oddi function after common bile duct stone management: evaluation from the viewpoint of endoscopic manometry, Gut, 41 (1997), 541–544. https://doi.org/10.1136/gut.41.4.541 doi: 10.1136/gut.41.4.541
|
| [27] |
J. A. Burger, A. Wiestner, Targeting B cell receptor signalling in cancer: preclinical and clinical advances, Nat. Rev. Cancer, 18 (2018), 148–167. https://doi.org/10.1038/nrc.2017.121 doi: 10.1038/nrc.2017.121
|
| [28] |
R. K. Martin, S. R. Damle, Y. A. Valentine, M. P. Zellner, B. N. James, J. C. Lownik, et al., B1 cell IgE impedes mast cell-mediated enhancement of parasite expulsion through B2 IgE blockade, Cell Rep., 22 (2018), 1824–1834. https://doi.org/10.1016/j.celrep.2018.01.048 doi: 10.1016/j.celrep.2018.01.048
|
| [29] |
S. Vallabhapurapu, M. Karin, Regulation and function of NF-kappaB transcription factors in the immune system, Annu. Rev. Immunol., 27 (2009), 693–733. https://doi.org/10.1146/annurev.immunol.021908.132641 doi: 10.1146/annurev.immunol.021908.132641
|
| [30] |
M. S. Hayden, S. Ghosh, Shared principles in NF-kappaB signaling, Cell, 132 (2008), 344–362. https://doi.org/10.1016/j.cell.2008.01.020 doi: 10.1016/j.cell.2008.01.020
|
| [31] |
S. C. Sun, Non-canonical NF-kappaB signaling pathway, Cell Res., 21 (2011), 71–85. https://doi.org/10.1038/cr.2010.177 doi: 10.1038/cr.2010.177
|
| [32] |
T. S. O. Jameson, G. F. Pavis, M. L. Dirks, B. P. Lee, D. R. Abdelrahman, A. J. Murton, et al., Reducing NF-kappaB signaling nutritionally is associated with expedited recovery of skeletal muscle function after damage, J. Clin. Endocrinol. Metab., 106 (2021), 2057–2076. https://doi.org/10.1210/clinem/dgab106 doi: 10.1210/clinem/dgab106
|
| [33] |
J. Choi, J. D. Phelan, G. W. Wright, B. Haupl, D. W. Huang, A. L. Shaffer, et al., Regulation of B cell receptor-dependent NF-kappaB signaling by the tumor suppressor KLHL14, Proc. Natl. Acad. Sci., 117 (2020), 6092–6102. https://doi.org/10.1073/pnas.1921187117 doi: 10.1073/pnas.1921187117
|
| [34] |
C. I. E. Smith, J. A. Burger, Resistance mutations to BTK inhibitors originate from the NF-kappaB but not from the PI3K-RAS-MAPK arm of the B cell receptor signaling pathway, Front. Immunol., 12 (2021), 689472. https://doi.org/10.3389/fimmu.2021.689472 doi: 10.3389/fimmu.2021.689472
|
| [35] |
R. Hurdayal, F. Brombacher, Interleukin-4 receptor alpha: From innate to adaptive immunity in murine models of cutaneous leishmaniasis, Front. Immunol., 8 (2017), 1354. https://doi.org/10.3389/fimmu.2017.01354 doi: 10.3389/fimmu.2017.01354
|
| [36] |
B. Guo, L. Zhang, N. Chiorazzi, T. L. Rothstein, IL-4 rescues surface IgM expression in chronic lymphocytic leukemia, Blood, 128 (2016), 553–562. https://doi.org/10.1182/blood-2015-11-682997 doi: 10.1182/blood-2015-11-682997
|
| [37] |
S. Freeley, C. Kemper, G. L. Friec, The "ins and outs" of complement-driven immune responses, Immunol. Rev., 274 (2016), 16–32. https://doi.org/10.1111/imr.12472 doi: 10.1111/imr.12472
|
| [38] |
K. K. Lee, Y. Omiya, M. Yuzurihara, Y. Kase, H. Kobayashi, Antispasmodic effect of shakuyakukanzoto extract on experimental muscle cramps in vivo: Role of the active constituents of Glycyrrhizae radix, J. Ethnopharmacol., 145 (2013), 286–293. https://doi.org/10.1016/j.jep.2012.11.005 doi: 10.1016/j.jep.2012.11.005
|
| [39] |
A. Madejon, J. Sheldon, I. Francisco-Recuero, C. Perales, M. Dominguez-Beato, M. Lasa, et al., Hepatitis C virus-mediated Aurora B kinase inhibition modulates inflammatory pathway and viral infectivity, J. Hepatol., 63 (2015), 312–319. https://doi.org/10.1016/j.jhep.2015.02.036 doi: 10.1016/j.jhep.2015.02.036
|
| [40] |
Z. Wang, Z. Yu, G. H. Wang, Y. M. Zhou, J. P. Deng, Y. Feng, et al., AURKB promotes the metastasis of gastric cancer, possibly by inducing EMT, Cancer Manage. Res., 12 (2020), 6947–6958. https://doi.org/10.2147/CMAR.S254250 doi: 10.2147/CMAR.S254250
|
| [41] |
M. Suelves, R. Lopez-Alemany, F. Lluis, G. Aniorte, E. Serrano, M. Parra, et al., Plasmin activity is required for myogenesis in vitro and skeletal muscle regeneration in vivo, Blood, 99 (2002), 2835–2844. https://doi.org/10.1182/blood.v99.8.2835 doi: 10.1182/blood.v99.8.2835
|
| [42] |
J. P. Vago, M. A. Sugimoto, K. M. Lima, G. L. Negreiros-Lima, N. Baik, M. M. Teixeira, et al., Plasminogen and the plasminogen receptor, Plg-RKT, regulate macrophage phenotypic, and functional changes, Front. Immunol., 10 (2019), 1458. https://doi.org/10.3389/fimmu.2019.01458 doi: 10.3389/fimmu.2019.01458
|
| [43] |
Y. Sato, T. Akao, J. X. He, H. Nojima, Y. Kuraishi, T. Morota, et al., Glycycoumarin from Glycyrrhizae Radix acts as a potent antispasmodic through inhibition of phosphodiesterase 3, J. Ethnopharmacol., 105 (2006), 409–414. https://doi.org/10.1016/j.jep.2005.11.017 doi: 10.1016/j.jep.2005.11.017
|
| [44] |
G. Xu, S. Fu, X. Zhan, Z. Wang, P. Zhang, W. Shi, et al., Echinatin effectively protects against NLRP3 inflammasome-driven diseases by targeting HSP90, JCI Insight, 6 (2021), e134601. https://doi.org/10.1172/jci.insight.134601 doi: 10.1172/jci.insight.134601
|
| [45] |
K. Yonekura-Sakakibara, Y. Higashi, R. Nakabayashi, The origin and evolution of plant flavonoid metabolism, Front. Plant. Sci., 10 (2019), 943. https://doi.org/10.3389/fpls.2019.00943 doi: 10.3389/fpls.2019.00943
|
| [46] |
B. Orlikova, M. Schnekenburger, M. Zloh, F. Golais, M. Diederich, D. Tasdemir, Natural chalcones as dual inhibitors of HDACs and NF-kappaB, Oncol. Rep., 28 (2012), 797–805. https://doi.org/10.3892/or.2012.1870 doi: 10.3892/or.2012.1870
|








Figures(9) / Tables(4)

Yong-hong Hu, Xue-ying Wang, Xi-wen Zhang, Jian Chen, Fu Li. Investigation of the mechanisms and experimental verification of Shao yao gan cao decoction against Sphincter of Oddi Dysfunction via systems pharmacology[J]. Mathematical Biosciences and Engineering, 2022, 19(12): 13374-13398. doi: 10.3934/mbe.2022626







 DownLoad:
DownLoad: