Prostate cancer is the fifth most common cause of death from cancer, and the second most common diagnosed cancer in men. In the last few years many mathematical models have been proposed to describe the dynamics of prostate cancer under treatment. So far one of the major challenges has been the development of mathematical models that would represent in vivo conditions and therefore be suitable for clinical applications, while being mathematically treatable. In this paper, we take a step in this direction, by proposing a nonlinear distributed-delay dynamical system that explores neuroendocrine transdifferentiation in human prostate cancer in vivo. Sufficient conditions for the existence and the stability of a tumour-present equilibrium are given, and the occurrence of a Hopf bifurcation is proven for a uniform delay distribution. Numerical simulations are provided to explore differences in behaviour for uniform and exponential delay distributions. The results suggest that the choice of the delay distribution is key in defining the dynamics of the system and in determining the conditions for the onset of oscillations following a switch in the stability of the tumour-present equilibrium.
Citation: Leo Turner, Andrew Burbanks, Marianna Cerasuolo. PCa dynamics with neuroendocrine differentiation and distributed delay[J]. Mathematical Biosciences and Engineering, 2021, 18(6): 8577-8602. doi: 10.3934/mbe.2021425
Prostate cancer is the fifth most common cause of death from cancer, and the second most common diagnosed cancer in men. In the last few years many mathematical models have been proposed to describe the dynamics of prostate cancer under treatment. So far one of the major challenges has been the development of mathematical models that would represent in vivo conditions and therefore be suitable for clinical applications, while being mathematically treatable. In this paper, we take a step in this direction, by proposing a nonlinear distributed-delay dynamical system that explores neuroendocrine transdifferentiation in human prostate cancer in vivo. Sufficient conditions for the existence and the stability of a tumour-present equilibrium are given, and the occurrence of a Hopf bifurcation is proven for a uniform delay distribution. Numerical simulations are provided to explore differences in behaviour for uniform and exponential delay distributions. The results suggest that the choice of the delay distribution is key in defining the dynamics of the system and in determining the conditions for the onset of oscillations following a switch in the stability of the tumour-present equilibrium.

| [1] |
F. Bray, J. Ren, E. Masuyer, J. Ferlay, Global estimates of cancer prevalence for 27 sites in the adult population in 2008, Int. J. Cancer, 132 (2012), 1133–1145. doi: 10.1002/ijc.27711. doi: 10.1002/ijc.27711

|
| [2] | W. H. Organisation, Cancer Today, available from: https://gco.iarc.fr/today, Last Accessed: 2020-06-05. |
| [3] |
P. J. Hensley, N. Kyprianou, Modeling prostate cancer in mice: limitations and opportunities, J. Androl., 33 (2012), 133–144. doi: 10.2164/jandrol.111.013987. doi: 10.2164/jandrol.111.013987
|
| [4] | J. Horoszewicz, S. Leong, T. Ming-Chu, Z. Wajsman, M. Friedman, L. Papsidero, et al., The LNCaP cell line - a new model for studies on human prostatic carcinoma, Prog. Clin. Biol. Res., 37 (1980), 115–132. |
| [5] |
T. Phan, S. Crook, A. Bryce, C. Maley, E. Kostelich, Y. Kuang, Mathematical modeling of prostate cancer and clinical application, Appl. Sci., 10 (2020), 2721. doi: 10.3390/app10082721. doi: 10.3390/app10082721
|
| [6] |
K. Swanson, L. True, D. Lin, K. Buhler, R. Vessella, J. Murray, A quantitative model for the dynamics of serum prostate-specific antigen as a marker for cancerous growth: An explanation for a medic anomaly, Am. J. Pathol., 163 (2001), 2513–2522. doi: 10.1016/S0002-9440(10)64691-3. doi: 10.1016/S0002-9440(10)64691-3
|
| [7] |
R. Vollmer, S. Egaqa, S. Kuwao, S. Baba, The dynamics of prostate antigen during watchful waiting of prostate carcinoma: A study of 94 japanese men, Cancer, 94 (2002), 1692–1698. doi: 10.1002/cncr.10443. doi: 10.1002/cncr.10443
|
| [8] |
R. Vollmer, P. Humphrey, Tumor volume in prostate cancer and serum prostate-specific antigen: Analysis from a kinetic viewpoint, Am. J. Pathol., 119 (2003), 80–89. doi: 10.1309/UNAQ-JTFP-B1RQ-BQD4. doi: 10.1309/UNAQ-JTFP-B1RQ-BQD4
|
| [9] |
Y. Kuang, J. Nagy, J. Elser, Biological stoichiometry of tumor dynamics: mathematical models and analysis, Disc. Cont. Dyn. Sys. B, 4 (2004), 221–240. doi: 10.3934/dcdsb.2004.4.221. doi: 10.3934/dcdsb.2004.4.221
|
| [10] |
C. Heinlein, C. Chang, Androgen receptor in prostate cancer, Endocr. Rev., 25 (2004), 276–308. doi: 10.1210/er.2002-0032. doi: 10.1210/er.2002-0032
|
| [11] | P. Koivisto, M. Kolmer, T. Visakorpi, O. Kallioniemi, Androgen receptor gene and hormonal therapy failure of prostate cancer, Am. J. Pathol., 152 (1998), 1–9. |
| [12] |
R. Rittmaster, A. Manning, A. Wright, L. Thomas, S. Whitefield, R. Norman, et al., Evidence for atrophy and apoptosis in the ventral prostate of rats given the 5 alpha-reductase inhibitor finasteride, Endocrinology, 136 (1995), 741–748. doi: 10.1210/endo.136.2.7835306. doi: 10.1210/endo.136.2.7835306
|
| [13] |
T. Jackson, A mathematical investigation of the multiple pathways to recurrent prostate cancer: Comparison with experimental data, Neoplasia, 6 (2004), 697–704. doi: 10.1593/neo.04259. doi: 10.1593/neo.04259
|
| [14] |
T. Jackson, A mathematical model of prostate tumor growth and androgen-independent relapse, Disc. Cont. Dyn. Sys. B, 4 (2004), 187–201. doi: 10.3934/dcdsb.2004.4.187. doi: 10.3934/dcdsb.2004.4.187
|
| [15] |
A. Ideta, G. Tanaka, T. Takeuchi, K. Aihara, A mathematical model of intermittent androgen suppression for prostate cancer, J. Nonlinear Sci., 18 (2008), 593–614. doi: 10.1007/s00332-008-9031-0. doi: 10.1007/s00332-008-9031-0
|
| [16] | S. Eikenberry, J. Nagy, Y. Kuang, The evolutionary impact of androgen levels on prostate cancer in a multi-scale mathematical model, Biol. Direct, 5. doi: 10.1186/1745-6150-5-24. |
| [17] |
S. Terry, H. Beltran, The many faces of neuroeondocrine differentiation in prostate cancer progression, Front. Oncol., 4 (2014), 1–9. doi: 10.3389/fonc.2014.00060. doi: 10.3389/fonc.2014.00060
|
| [18] |
V. Perrot, Neuroendocrine differentiation in the progression of prostate cancer: an update to recent developments, Open J. Urol., 2 (2012), 173–182. doi: 10.4236/oju.2012.223032. doi: 10.4236/oju.2012.223032
|
| [19] |
C. Shen, Z. Burke, D. Tosh, Transdifferentiation, metaplasia and tissue regeneration, Organogenesis, 1 (2004), 36–44. doi: 10.4161/org.1.2.1409. doi: 10.4161/org.1.2.1409
|
| [20] |
M. Cerasuolo, D. Paris, F. A. Iannotti, D. Melck, R. Verde, E. Mazzarella, A. Motta, A. Ligresti, Neuroendocrine transdifferentiation in human prostate cancer cells: an integrated approach, Cancer Res., 75 (2015), 2975–2986. doi: 10.1158/0008-5472.CAN-14-3830. doi: 10.1158/0008-5472.CAN-14-3830
|
| [21] |
M. Adimy, F. Crauste, C. Marquet, Asymptotic behaviour and stability switch for a mature-immature model of cell differentiation, Nonlinear Anal. Real World Appl., 11 (2010), 2913–2929. doi: 10.1016/j.nonrwa.2009.11.001. doi: 10.1016/j.nonrwa.2009.11.001
|
| [22] |
M. Adimy, F. Crauste, S. Ruan, Modelling hematopoiesis mediated by growth factors with applications to periodic hematological diseases, Bull. Math. Biol., 68 (2006), 2321–2351. doi: 10.1007/s11538-006-9121-9. doi: 10.1007/s11538-006-9121-9
|
| [23] |
J. Morken, A. Packer, R. Everett, J. Nagy, Y. Kuang, Mechanisms of resistance to intermittent androgen deprivation in patients with prostate cancer identified by a novel computational method, Cancer Res., 74 (2014), 3673–3683. doi: 10.1158/0008-5472.CAN-13-3162. doi: 10.1158/0008-5472.CAN-13-3162
|
| [24] |
L. Turner, A. Burbanks, M. Cerasuolo, Mathematical insights into neuroendocrine transdifferentiation of human prostate cancer cells, Nonlinear Anal. Model. Control, 26 (2021), 884–913. doi: 10.15388/namc.2021.26.24441. doi: 10.15388/namc.2021.26.24441
|
| [25] |
G. Hutchinson, Circular causal systems in ecology., Ann. N. Y. Acad. Sci., 50 (1948), 221–246. doi: 10.1111/j.1749-6632.1948.tb39854.x. doi: 10.1111/j.1749-6632.1948.tb39854.x
|
| [26] |
T. Cassidy, A. R. Humphries, A mathematical model of viral oncology as an immuno-oncology instigator, Math. Med. Biol., 37 (2020), 117–151. doi: 10.1093/imammb/dqz008. doi: 10.1093/imammb/dqz008
|
| [27] | F. Scudo, J. Ziegler, The golden age of theoretical ecology, 1923-1940: A collection of the works of V.Volterra, V.A. Kostitzin, A.J. Lotka, and A.N. Kolmogoroff, vol. 22, Springer, 1978. |
| [28] |
J. Baez, Y. Kuang, Mathematical models of androgen resistance in prostate cancer patients under intermittent androgen suppression therapy, Appl. Sci., 6 (2016), 352. doi: 10.3390/app6110352. doi: 10.3390/app6110352
|
| [29] |
M. Droop, Vitamin B12 and marine ecology, IV: The kinetics of uptake, growth and inhibition in Monochrysis lutheri, J. Mar. Biol. Assoc, UK, 48 (1968), 689–733. doi: 10.1017/S0025315400019238. doi: 10.1017/S0025315400019238
|
| [30] |
B. Buonomo, M. Cerasuolo, The effect of time delay in plant-pathogen interactions with host demography, Math. Biosci. Eng., 12 (2015), 473–490. doi: 10.3934/mbe.2015.12.473. doi: 10.3934/mbe.2015.12.473
|
| [31] |
T. Faria, E. Liz, Boundedness and asymptotic stability for delayed equations of logistic type., Proc. Math. Roy. Soc. Edinb., 133 (2003), 1057–1073. doi: 10.1017/S030821050000281X. doi: 10.1017/S030821050000281X
|
| [32] |
A. Ahmadian, M. Bin Suleiman, F. Ismail, Numerical simulation of tumor development stages using artificial neural network., Trends Appl. Sci. Res., 7 (2012), 132–141. doi: 10.3923/tasr.2012.132.141. doi: 10.3923/tasr.2012.132.141
|
| [33] |
M. Piotrowska, M. Bodnar, Influence of distributed delays on the dynamics of a generalized immune system cancerous cells interactions model, Commun. Nonlinear Sci. Numer. Simulat., 54 (2018), 38. doi: 10.1016/j.cnsns.2017.06.003. doi: 10.1016/j.cnsns.2017.06.003
|
| [34] |
E. Beretta, Y. Kuang, Geometric stability switch criteria in delay differential systems with delay dependent parameters, SIAM J. Math. Anal., 33 (2002), 1144–1165. doi: 10.1137/S0036141000376086. doi: 10.1137/S0036141000376086
|
| [35] | H. Smith, An Introduction to Delay Differential Equations with Applications to the Life Sciences, Springer, New York, 2010. doi: 10.1007/978-1-4419-7646-8. |
| [36] |
P.-L. Clermont, X. Ci, H. Pandha, Y. Wang, F. Crea, Treatment-emergent neuroendocrine prostate cancer: molecularly driven clinical guidelines, Int. J. Endocr. Oncol., 6 (2019), IJE20. doi: 10.2217/ije-2019-0008. doi: 10.2217/ije-2019-0008
|
| [37] |
B. Rahman, K. Blyuss, Y. Kyrychko, Dynamics of neural systems with discrete and distributed time delays, SIAM J. Appl. Dyn., 14 (2015), 2069–2095. doi: 10.1137/15M1006398. doi: 10.1137/15M1006398
|
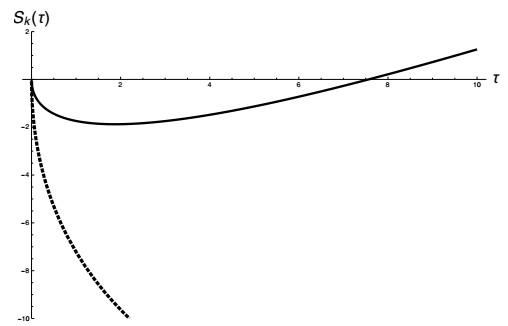
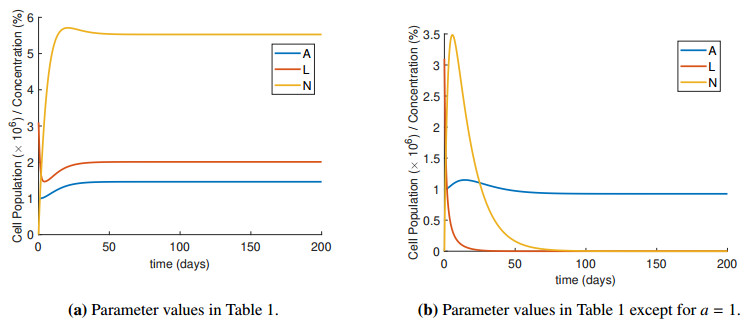
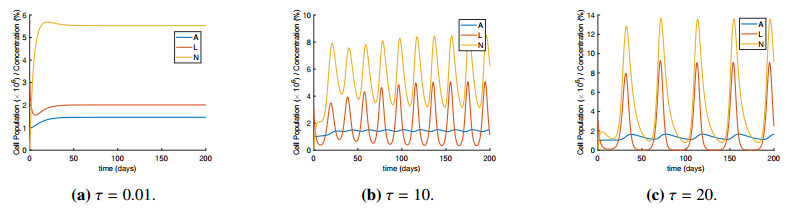
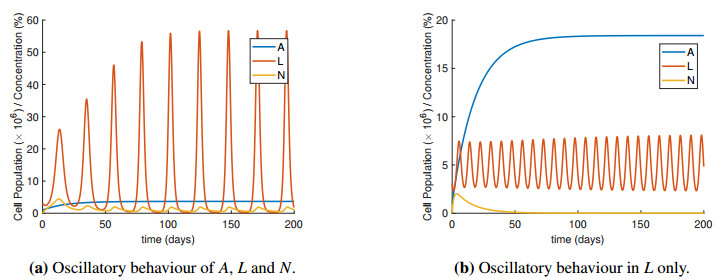
Figures(4) / Tables(2)

Leo Turner, Andrew Burbanks, Marianna Cerasuolo. PCa dynamics with neuroendocrine differentiation and distributed delay[J]. Mathematical Biosciences and Engineering, 2021, 18(6): 8577-8602. doi: 10.3934/mbe.2021425







 DownLoad:
DownLoad: