In this paper, we propose a prey-predator model with age structure which is described by the mature period. The aim of this paper is to study how mature period affect the dynamics of interaction between prey and predator. The sufficient condition of the existence of non-negative steady state is derived. By using integrated semigroup theory, we obtain the characteristic equation, by which we find that the non-negative steady state will lose its stability via Hopf bifurcation induced by mature period, and the corresponding periodic solutions emerge. Additionally, some numerical simulations are provided to illustrate the results predicted by linear analysis. Especially, the numerical results indicate that both mature period and age can affect the amplitude and period of periodic solutions.
Citation: Lijun Wang, Chuanjun Dai, Min Zhao. Hopf bifurcation in an age-structured prey-predator model with Holling Ⅲ response function[J]. Mathematical Biosciences and Engineering, 2021, 18(4): 3144-3159. doi: 10.3934/mbe.2021156
In this paper, we propose a prey-predator model with age structure which is described by the mature period. The aim of this paper is to study how mature period affect the dynamics of interaction between prey and predator. The sufficient condition of the existence of non-negative steady state is derived. By using integrated semigroup theory, we obtain the characteristic equation, by which we find that the non-negative steady state will lose its stability via Hopf bifurcation induced by mature period, and the corresponding periodic solutions emerge. Additionally, some numerical simulations are provided to illustrate the results predicted by linear analysis. Especially, the numerical results indicate that both mature period and age can affect the amplitude and period of periodic solutions.

| [1] |
M. L. Quammen, Predation by Shorebirds, Fish, and Crabs on Invertebrates in Intertidal Mudflats: An Experimental Test, Ecology, 65 (1984), 529–537. doi: 10.2307/1941415

|
| [2] |
H. D. Rundle, S. M. Vamosi, Experimental test of predation's effect on divergent selection during character displacement in sticklebacks, Proc. Natl. Acad. Sci. U.S.A., 100 (2003), 14943–14948. doi: 10.1073/pnas.2036360100
|
| [3] | K. G. Magn$\acute{u}$sson, O. K. P$\acute{a}$lsson, Predator–prey interactions of cod and capelin in Icelandic waters, Proc. Natl. Acad. Sci. U.S.A., 193 (1991), 153–170. |
| [4] |
J. H. Steele, E. W. Henderson, The role of predation in plankton models, J. Plankton Res., 14 (1992), 157–172. doi: 10.1093/plankt/14.1.157
|
| [5] |
R. Mohn, W. D. Bowen, Grey seal predation on the eastern Scotian Shelf: Modeling the impact on Atlantic cod, Can. J. Fish. Aquat. Sci., 53 (1996), 2722–2738. doi: 10.1139/f96-239
|
| [6] |
L. J. White, R. J. Maude, W. Pongtavornpinyo, S. Saralamba, R. Aguas, T. V. Effelterre, et al., The role of simple mathematical models in malaria elimination strategy design, Malar. J., 8 (2009), 212. doi: 10.1186/1475-2875-8-212
|
| [7] | V. Volterra, The role of simple mathematical models in malaria elimination strategy design, Roma:Academia Nazionale dei Lincei., 2 (1926), 31–113. |
| [8] | A. J. Lotka, Elements of physical biology, Science Progress in the Twentieth Century (1919–1933), 21 (1926), 341–343. |
| [9] |
W. Y. Wang, L. J. Pei, Stability and Hopf bifurcation of a delayed ratio-dependent predator-prey system, Acta Mech. Sin., 27 (2011), 285–296. doi: 10.1007/s10409-011-0424-1
|
| [10] |
J. H. Bao, J. H. Shao, Permanence and Extinction of Regime-Switching Predator-Prey Models, SIAM J. Math. Anal., 48 (2016), 725–739. doi: 10.1137/15M1024512
|
| [11] | C. S. Holling, The functional response of predators to prey density and its role in mimicry and population regulation, Mem. Ent. Soc. Can., 97 (1965), 5–60. |
| [12] |
J. P. Chen, Z. H. De, The qualitative analysis of two species predator-prey model with Holling's type Ⅲ functional response, Appl. Math. Mech., 7 (1986), 77–86. doi: 10.1007/BF01896254
|
| [13] | Y. J. Huang, F. D. Chen, L. Zhong, Stability analysis of a prey–predator model with Holling type Ⅲ response function incorporating a prey refuge, Appl. Math. Comput., 182 (206), 672–683. |
| [14] |
J. H. Wang, L. Q. Pan, Qualitative analysis of a harvested predator-prey system with Holling-type Ⅲ functional response incorporating a prey refuge, Adv. Differ. Equ., 2012 (2012), 96. doi: 10.1186/1687-1847-2012-96
|
| [15] |
R. Z. Yang, J. J. Wei, Stability and bifurcation analysis of a diffusive prey–predator system in Holling type Ⅲ with a prey refuge, Nonlinear Dyn., 79 (2015), 631–646. doi: 10.1007/s11071-014-1691-8
|
| [16] |
J. F. Wang, Spatiotemporal patterns of a homogeneous diffusive predator–prey system with Holling type Ⅲ functional response, J. Dyn. Differ. Equ., 29 (2017), 1383–1409. doi: 10.1007/s10884-016-9517-7
|
| [17] |
N. Apreutesei, G. Dimitriu, On a prey–predator reaction–diffusion system with Holling type Ⅲ functional response, J. Comput. Appl. Math., 235 (2010), 366–379. doi: 10.1016/j.cam.2010.05.040
|
| [18] |
P. J. Pal, P.K. Mandal, K. K. Lahiri, A delayed ratio-dependent predator–prey model of interacting populations with Holling type Ⅲ functional response, Nonlinear Dyn., 76 (2014), 201–220. doi: 10.1007/s11071-013-1121-3
|
| [19] | X. D. Wang, M. Peng, Stability and Hopf bifurcation analysis of a ratio-dependent predator–prey model with two time delays and Holling type Ⅲ functional response, Appl. Math. Comput., 268 (2015), 496–508. |
| [20] | Z. W. Cai, L. H. Huang, H. B. Chen, Positive periodic solution for a multispecies competition-predator system with Holling Ⅲ functional response and time delays, Appl. Math. Comput., 217 (2011), 4866–4878. |
| [21] |
E. G. Olivares, A. R. Palma, Multiple limit cycles in a Gause type predator–prey model with Holling type Ⅲ functional response and Allee effect on prey, Bull. Math. Biol., 73 (2011), 1378–1397. doi: 10.1007/s11538-010-9577-5
|
| [22] |
H. Liu, K. Zhang, Y. Ye, Y. M. Wei, M. Ma, Dynamic complexity and bifurcation analysis of a host–parasitoid model with Allee effect and Holling type Ⅲ functional response, Adv. Differ. Equ., 2019 (2019), 507. doi: 10.1186/s13662-019-2430-8
|
| [23] | S. Sengupta, P. Das, D. Mukherjee, Stochastic non-autonomous Holling type-Ⅲ prey-predator model with predator's intra-specific competition, Discrete Contin. Dyn. Syst., 23 (2018), 3275. |
| [24] |
X. M. Fan, F. Q. Jiang, H. N. Zhang, Dynamics of multi-species competition–predator system with impulsive perturbations and Holling type Ⅲ functional responses, Nonlinear Anal. Theory Methods Appl., 74 (2011), 3363–3378. doi: 10.1016/j.na.2011.02.012
|
| [25] |
D. J. Thompson, Towards a predator-prey model incorporating age structure: the effects of predator and prey size on the predation of Daphnia magna by Ischnura elegans, J. Anim. Ecol., 44 (1975), 907–916. doi: 10.2307/3727
|
| [26] |
E. M. Cauley, W. G. Wilson, Dynamics of age-structured and spatially structured predator-prey interactions: individual-based models and population-level formulations, Am. Nat., 142 (1993), 412–442. doi: 10.1086/285547
|
| [27] |
M. E. Gurtin, D. S. Levine, On predator-prey interactions with predation dependent on age of prey, Math. Biosci., 47 (1979), 207–219. doi: 10.1016/0025-5564(79)90038-5
|
| [28] | J. M. Cushing, M. Saleem, A competition model with age structure, in Mathematical Ecology, Springer, Berlin (1984), 178–192. |
| [29] | J. M. Cushing, M. Saleem, competition with age structure, in Population Biology, Springer, Berlin (1983), 225–230. |
| [30] |
J. M. Cushing, M. Saleem, A predator prey model with age structure, J. Math. Biol., 14 (1982), 231–250. doi: 10.1007/BF01832847
|
| [31] |
S. X. Zhang, X. X. Xu, A mathematical model for hepatitis B with infection-age structure, Discrete Contin. Dyn. Syst., 21 (2016), 1329–1346. doi: 10.3934/dcdsb.2016.21.1329
|
| [32] |
M. Saleem, Egg-eating age-structured predators in interaction with age-structured prey, Math. Biosci., 70 (1984), 91–104. doi: 10.1016/0025-5564(84)90048-8
|
| [33] | J. M. Cushing, An introduction to structured population dynamics, Society for industrial and applied mathematics, 1998. |
| [34] |
E. Beninc$\grave{a}$, J. Huisman, R. Heerkloss, K. D. J$\ddot{o}$hnk, P. Branco, E. H. V. Nes, et al., Chaos in a long-term experiment with a plankton community, Nature, 451 (2008), 822–825. doi: 10.1038/nature06512
|
| [35] |
E. Beninc$\grave{a}$, B. Ballantine, S. P. Ellner, J. Huisman, Species fluctuations sustained by a cyclic succession at the edge of chaos, Proc. Natl. Acad. Sci. U.S.A., 112 (2015), 6389–6394. doi: 10.1073/pnas.1421968112
|
| [36] |
J. A. Sherratt, M. J. Smith, Periodic travelling waves in cyclic populations: field studies and reaction–diffusion models, J. R. Soc. Interface, 5 (2008), 483–505. doi: 10.1098/rsif.2007.1327
|
| [37] |
Z. H. Liu, N. W. Li, Stability and bifurcation in a predator–prey model with age structure and delays, J. Nonlinear Sci., 25 (2015), 937–957. doi: 10.1007/s00332-015-9245-x
|
| [38] | X. M. Zhang, Z. H. Liu, Hopf bifurcation analysis in a predator-prey model with predator-age structure and predator-prey reaction time delay, Appl. Math. Model., 91 (2020), 530–548. |
| [39] |
X. M. Zhang, Z. H. Liu, Periodic oscillations in age-structured ratio-dependent predator–prey model with Michaelis–Menten type functional response, Physica D, 389 (2019), 51–63. doi: 10.1016/j.physd.2018.10.002
|
| [40] |
H. Tang, Z. H. Liu, Hopf bifurcation for a predator–prey model with age structure, Appl. Math. Model., 40 (2016), 726–737. doi: 10.1016/j.apm.2015.09.015
|
| [41] | P. Magal, S. G. Ruan, On semilinear Cauchy problems with non-dense domain, Adv. Differ. Equ., 14 (2009), 1041–1084. |
| [42] | P. Magal, Compact attractors for time-periodic age-structured population models, Electron. J. Differ. Equ., 2001 (2001), 1–35. |
| [43] |
Z. H. Liu, P. Magal, S. G. Ruan, Hopf bifurcation for non-densely defined Cauchy problems, Z. Angew. Math. Phys., 62 (2011), 191–222. doi: 10.1007/s00033-010-0088-x
|
| [44] |
W. Zhen, Z. H. Liu, Hopf bifurcation of an age-structured compartmental pest-pathogen model, J. Math. Anal. Appl., 385 (2012), 1134–1150. doi: 10.1016/j.jmaa.2011.07.038
|
| [45] |
L. M. Abia, O. Angulo, J. C. L$\acute{o}$pez-Marcos, Age-structured population models and their numerical solution, Ecol. Modell., 188 (2005), 112–136. doi: 10.1016/j.ecolmodel.2005.05.007
|
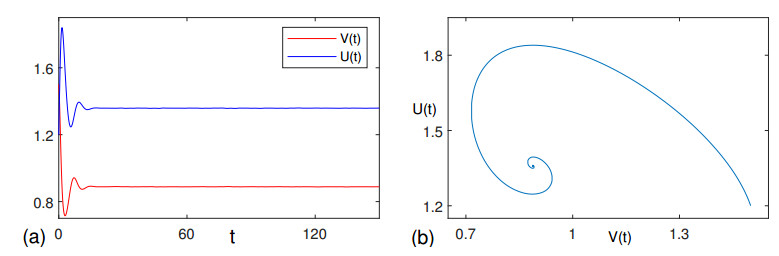



Figures(4)

Lijun Wang, Chuanjun Dai, Min Zhao. Hopf bifurcation in an age-structured prey-predator model with Holling Ⅲ response function[J]. Mathematical Biosciences and Engineering, 2021, 18(4): 3144-3159. doi: 10.3934/mbe.2021156







 DownLoad:
DownLoad: